
Запрещённые пептиды в профессиональном спорте
23.05.2020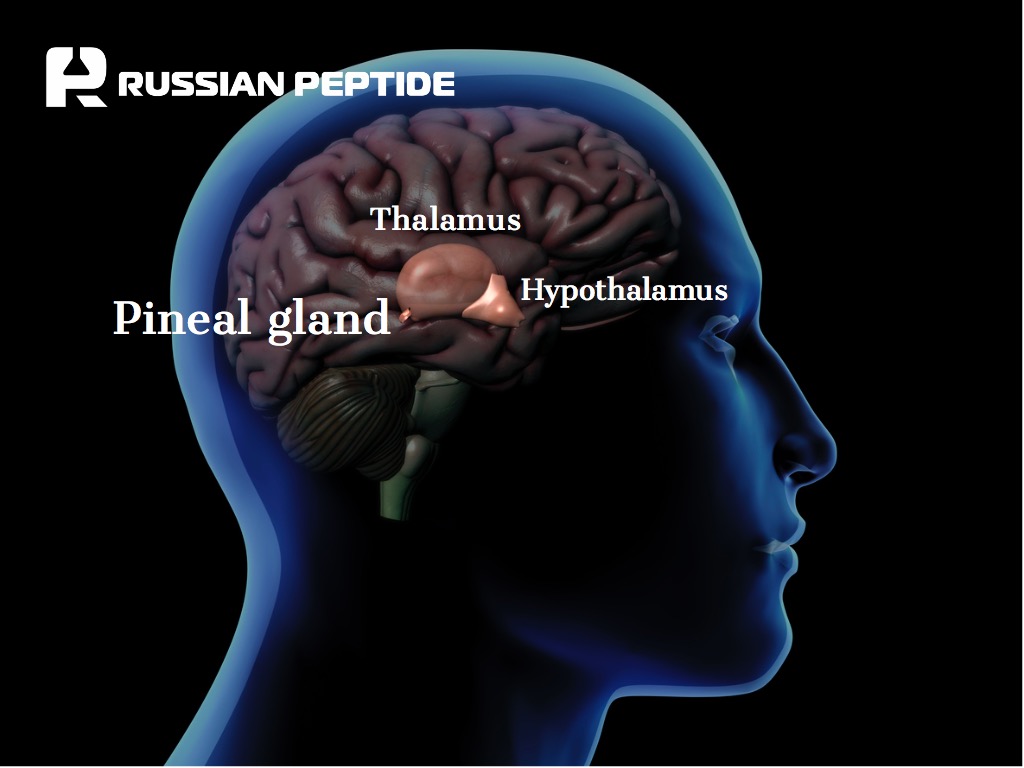
Пептиды шишковидной железы
04.06.2020
Миостатин, ингибиторы миостатина, фоллистатин 344 и 315, мышцы, допинг, геронтология
Нарушения состава тканей организма могут возникнуть при любом весе тела. Измерения массы тела и индекс массы тела долгое время считались практичными и подходящими параметрами для прогнозирования рисков для здоровья человека. Низкая мышечная масса является предиктором заболеваемости и смертности, будучи связанной с такими последствиями, как повышение риска хирургических и послеоперационных осложнений, увеличение продолжительности пребывания в стационаре, снижение физических функций, ухудшение качества жизни и сокращение ее продолжительности [1]. Причинами таких нарушений могут быть рак, болезни печени, почек, поджелудочной железы, сердечно-сосудистые нарушения, ХОБЛ, аспирационная пневмония, грыжи брюшной стенки, инфекционные заболевания и старение, с которым зачастую связана саркопения – атрофическое дегенеративное изменение скелетной мускулатуры, ведущее к потере силы и мышечной массы. В целом, более высокая мышечная масса обычно связана с лучшими прогнозами, включая продолжительность жизни [1].
Одним из регуляторов роста мышц является белок миостатин (Myostatin), также называемый «фактор роста и дифференцировки-8» (GDF-8), из суперсемейства белков трансформирующего фактора роста-β (TGF-β). Миостатин первоначально был охарактеризован в 1997 году как специфический регулятор массы скелетных мышц у мышей [2]. Активная форма миостатина массой 12,5 кДа (107 АА <аминокислотных остатков>) образуется в результате внутриклеточного процессинга белка-предшественника массой 47,5 кДа (375 АА), который содержит сигнальную последовательность (АА1–23), присоединенную к пропептиду массой 35 кДа (АА 24–266), и активный фрагмент массой 12,5 кДа (AA267–375). Первичная структура миостатина содержит девять цистеиновых остатков, что позволяет образовывать четыре внутримолекулярные дисульфидные связи с одним свободным остатком цистеина. Общий заряд миостатина (pI = 6,4) показывает, что он является нейтральным пептидом. В системе кровообращения миостатин обнаруживается в связанном состоянии с про-пептидом [3].
Миостатин экспрессируется преимущественно в скелетных мышцах. На молекулярном уровне показано, что миостатин регулирует рост мышц не только путем контроля пролиферации и дифференцировки миобластов во время миогенеза плода, но также с помощью активации сателлитных клеток в постнатальном периоде [2]. Сигнальный путь миостатина важен для регуляции роста скелетных мышц. Как следует из названия, миостатин участвует в торможении роста мышц, противодействуя пути mTOR. У здорового человека существует баланс между обоими путями, что обеспечивает нормальный рост мышц и защищает от неконтролируемой клеточной пролиферации. Как локальная сигнальная молекула, миостатин инициирует расщепление белка посредством его связывания с мембраносвязанным рецептором на скелетных мышцах [4]. Встречающиеся в природе мутации в гене-продуценте миостатина быка, овцы, собаки и человека или инактивация гена у мышей приводят к увеличению мышечной массы вследствие гиперплазии [2, 5]. В ряде исследований было показано, что инъекции блокаторов миостатина, включая фоллистатин, антитела к миостатину и продомен домена миостатина, независимо друг от друга увеличивают регенерацию и рост мышц на мышиной модели мышечной дистрофии. Кроме того, длительное отсутствие миостатина у мышей также снижает потерю саркопенических мышц благодаря эффективной активации сателлитных клеток и регенерации скелетных мышц у старых мышей. Аналогичные эксперименты на стареющих мышах Mstn-ant1 показали увеличение активации сателлитных клеток и повышение эффективности регенерации мышц. Было установлено, что усиленный мышечный фенотип у мышей с нулевой выработкой миостатина приводит к сочетанию мышечной гиперплазии и гипертрофии (Рис. 1; адаптирован из работы [5]).

Рисунок 1 – Беспородная мышь (слева) и нуль-миостатиновая мышь (справа)
Мутантные нуль-миостатиновые мыши по отношению к беспородным животным имели массу мышц тела 135%, двубрюшная – 205%, грудная – 262%, трицепс плечевой – 235%, квадрицепс – 203%, икроножная/подошвенная – 219%, Tibialis cranialis – 202% и Soleus – 200% [5]. Как видно из приведенных значений, массы мышц практически удвоились, что привело к появлению в англоязычной научной литературе термина «double muscling» – «удвоение мышц».
Было выявлено несколько встречающихся в природе мутаций у различных пород крупного рогатого скота, таких как бельгийская голубая и Пьемонтезе (Piemontese). Мутации в кодирующей области гена миостатина приводит к экспрессии нефункционального белкового продукта, следствием чего является гипертрофия мышц животного [2] (Рисунок 2).

Рисунок 2 – Корова мясной породы (слева) и бык с подавленной выработкой миостатина (справа)
Также известно, что мутация в гене миостатина приводит к резко выраженной гипертрофии у детей [6]. Учитывая, что ингибирование миостатина приводит к значительному увеличению постнатального роста мышц, антагонисты миостатина имеют огромное терапевтическое значение для уменьшения потери мышечной массы. Дефицит миостатина проявляется в гипертрофии мышц и повышенной физической работоспособности [4], поэтому ген миостатина находится в центре пристального внимания в отношении генной терапии и допинга.
Миостатин активируется при болезненных состояниях и во время длительного постельного режима, когда мышечная недостаточность является симптоматической. Также миостатин присутствует в относительно высоких концентрациях в мышцах стареющих людей [4]. Отсюда следует, что ингибирование миостатина имело бы огромный потенциал для укрепления здоровья. Клинические применения для разработки лекарств на основе ингибиторов миостатина включают мышечную дистрофию, кахексию (мышечную атрофию, связанную со СПИДом и другими хроническими заболеваниями), миопатии, возникающие в результате воспаления, и саркопении, потерю мышечной массы, зачастую связанную с увеличением возраста. Тем не менее, лекарства, которые управляют передачей сигналов миостатина, в настоящее время рассматриваются в качестве препаратов для антивозрастной терапии и их потенциал используется для повышения физической работоспособности [7]. Фоллистатин, по-видимому, является привлекательным для будущего применения терапевтическим средством, которое блокирует активацию сигнального пути миостатина [8].
Фоллистатин (Follistatin, FS), как и другие члены суперсемейства TGF-β, например, активины и BMP-2, также играет роль в регуляции массы скелетных мышц [9]. Фоллистатин представляет собой белок массой 35 кДа (288 АА), выступающий в качестве важного медиатора секреции, развития и дифференцировки клеток в ряде систем тканей и органов. В 1987 году он был впервые выделен из фолликулярной жидкости яичника в качестве белкового фактора, способного подавлять секрецию фоллитропина клетками гипофиза в культуре [10]. Позже выяснилось, что этот эффект является лишь одним из многих его свойств в репродуктивных и непродуктивных системах. FS регулирует клеточную дифференцировку и секрецию благодаря своей мощной способности связывать и бионейтрализовать активин, с которым он локализуется во многих тканевых системах [10]. Фоллистатин является секретируемым гликопротеином, который содержится во многих тканях и органах (например, гипофиз, костный мозг, яичник, почка, печень, кровеносные сосуды). Типичные концентрации в сыворотке человека находятся в диапазоне 2–10 нг/мл. У женщин в норме 0,6–2,8 нг/мл, в зависимости от периода менструального цикла или беременности; у мужчин – 0,45 нг/мл. Фоллистатин-315 является преобладающей формой в кровообращении человека [11]. Из-за альтернативного сплайсинга гена FS у человека существуют две основные изоформы FS, а именно «FS315» («FS344») и «FS288» («FS317») [12]. Названия «FS315» и «FS288» относятся к общему количеству аминокислот, а альтернативные «FS344» и «FS317» – к последней аминокислоте непроцессированного белка, который состоит из 344 аминокислот [13]. Однако после трансляцииN-концевой сигнальный пептид, содержащий первые 29 аминокислот, отщепляется (Табл. 1). В то время как FS315 является основной изоформой, циркулирующей в крови, FS288 в основном обнаруживается в тканях.
Протеолитическое расщепление на С-конце FS315 может привести к дальнейшим изоформам, то есть FS300 (FS329) и FS303 (FS332). Белок содержит два сайта N—гликозилирования (N124, N288) и большое количество цистеинов (36), которые связаны друг с другом через внутримолекулярные дисульфидные мостики [13].
Таблица 1 – Характеристика аминокислотной последовательности человеческого фоллистатина* [12]
| 1–50 | MVRARHQPGG | LCLLLLLLCQ | FMEDRSAQA29G | NCWLRQAKNG | RCQVLYKTEL |
| 51–100 | SKEECCSTGR | LSTSWTEEDV | NDNTLFKWMI | FNGGAPNCIP | CKETCENVDC |
| 101–150 | GPGKKCRMNK | KNKPRCVCAP | DCSN124ITWKGP | VCGLDGKTYR | NECALLKARC |
| 151–200 | KEQPELEVQY | QGRCKKTCRD | VFCPGSSTCV | VDQTNNAYCV | TCNRICPEPA |
| 201–250 | SSEQYLCGND | GVTYSSACHL | RKATCLLGRS | IGLAYEGKCI | KAKSCEDIQC |
| 251–300 | TGGKKCLWDF | KVGRGRCSLC | DELCPDSKSD | EPVCASDN288AT | YASECAMKEA |
| 301–344 | ACSSGVLLEV | KHSGSCN317SIS | EDTEEEEED329E | DQ332DYSFPISS | ILEW344 |
* После трансляции первые 29 аминокислот (сигнальный пептид). Два сайта N-гликозилирования обнаружены на Asn124 и Asn288. Из-за альтернативного сплайсинга и посттрансляционной обработки существует несколько изоформ: FS288 (317), FS315 (344), FS300 (329), FS303 (332). Таким образом, везде, где специально не указан номер, под фоллистатином понимается изоформа FS315 (FS344).
Рекомбинантные человеческие фоллистатины для исследовательских целей получают экспрессированием в прокариотических (E.coli) и эукариотических клетках насекомых (Sf9, Sf21), яичниках китайского хомячка (CHO), эмбриональных клетках почки человека (HEK293) [13, 14]. Необходимая последовательность изоформы фоллистатина может быть получена с помощью ПЦР-амплификации соответствующего фрагмента мРНК из тканей человека (или других видов) и вставкой кДНК в вектор экспрессии млекопитающего, такой как плазмида, вирус или фаг, который может быть трансфицирован в клетку-продуцент для экспрессии требуемого пептида [15].
Фоллистатин связывается с белками семейства TGF-β (например, BMPs, GDF9, GDF8) и может нейтрализовать их функции. Миостатин, как описано выше, ингибирует рост и дифференцировку мышечных клеток (миогенез). Таким образом, ингибирование миостатина фоллистатином приводит к ускоренному развитию мышечной массы.
До настоящего времени большинство исследований по введению фоллистатина проводилось на животных моделях. Недавно были завершены клинические испытания Фазы 1 и 2 по переносу гена фоллистатина rAAV1.CMV.huFollistatin344 пациентам с мышечной дистрофией Беккера и спорадическим включением миозита тела (NCT01519349) и по внутримышечной передачи гена пациентам с мышечной дистрофией Дюшенна (NCT02354781), проведенные на базе Исследовательского центра Nationwide Children’s Hospital (Колумбус, штат Огайо, США) в рамках изучения подходов лечения с использованием генной терапии [16]. Обширные доклинические исследования с использованием аденоассоциированного вируса (AAV) для доставки фоллистатина на животных моделях (нормальные и дистрофические мыши mdx) продемонстрировали увеличение силы и размера мышц во всех случаях и без побочных эффектов [17]. Исследователи применили однократное введение гена белков-ингибиторов миостатина (AAV1-FS) нормальным и дистрофическим мышам в дозировке 1×1011 (высокая) и 1×1010 (низкая) вирусных частиц. Эксперимент длился в течение >2 лет, и положительный результат наблюдался даже у пожилых животных. Помимо фоллистатина были введены продуценты других белков, которые участвуют в регуляции миостатина: фоллистатин-родственный ген (FLRG), который in vitro ингибирует активин и костные морфогенетические белки и сывороточный фактор роста и дифференцировки (GASP-1), который, содержит несколько доменов, связанных с белками-ингибиторами протеазы, и домен, гомологичный повтору 10-цистеина, обнаруженному в фоллистатине, и непосредственно связывается со зрелым миостатином и пропилептидом миостатина, ингибиря активность миостатина. Их вводили с помощью аналогичных векторов – AAV1-GASP-1, AAV1-FLRG, и AAV1-GFP (кодирует зелёный флуоресцентный белок; контрольная группа). Наилучшие результаты показало введения AAV1-FS и вызванная им выработка фоллистатина. Через 2 года масса тела мышей составляла ~180% от таковой в контрольной группе, масса Tibialis cranialis удвоилась, а икроножной мышцы и квадрицепса – увеличилась почти в три раза [17]. Кроме того, сила мышц, в среднем, увеличилась на 30–40% на 180 день эксперимента, а также наблюдалось снижение уровней маркеров мышечного повреждения и старения.
Аналогичные эффекты были подтверждены на приматах [18]. Введение вектора AAV1-FS344 индуцировало выраженное и длительное увеличение размера и силы мышц у обезьян вида яванский макак (Macaca fascicularis). Шести животным ввели по 3×500 мкл раствора, содержащего в сумме 1×1013 вирусных частиц. Отметим, что человеческий FS344 имеет 98% гомологию с фоллистатином у нечеловекообразных приматов, поэтому животным дополнительно назначили иммунодепрессанты такролимус (2,0 мг/кг) и мофетил микофенилат (50 мг/кг) за 2 недели до введения вектора. Макаки в группе FS показали 15% увеличение окружности четырехглавой мышцы по сравнению с исходным уровнем через 8 недель после введения вектора, затем последовала стабилизация роста в течение 20-ой недели. Гистологические срезы четырехглавой мышцы через 20 недель показали увеличение среднего диаметра волокна в обработанной группе по сравниванию с контрольной на 34%. Измерения силы подергивания в обработанных мышцах двух макак показали увеличение на 11,8% и 35,7%. Эти же животные имели увеличение тетанической силы на 12,3% и 77,9% по сравнению с измерениями на необработанной стороне. Этот результат может указывать на то, что увеличение размера мышц после повышения уровня фоллистатина не обязательно приводит к пропорциональному возрастанию силы, как это было видно с другими подходами к ингибированию миостатина

Опираясь на доклинические результаты, в исследовании NCT01519349 авторы использовали альтернативно сплайсированный фоллистатин-344, чтобы избежать потенциального связывания с нецелевыми сайтами. AAV1.CMV.FS344 был доставлен шести пациентам с мышечной дистрофией Беккера путем прямых внутримышечных инъекций в четырехглавую мышцу с двух сторон. Группа 1 включала трех субъектов в возрасте 34±3,6 лет, получавших 3×1011 вг/кг/нога. Основным критерием оценки было пройденное расстояние в тесте с шестиминутной ходьбой (6MWT). Пациенты 01 и 02 улучшили показатели на 58 и 125 метров, соответственно. Пациент 03 не показал изменений. В когорте 2 (возраст 29±5,0 лет) пациенты 05 и 06 получали 6×1011 вг/кг/нога и показали улучшения в тесте 6MWT на 108 м и 29 м, тогда как у пациента 04 прогресса не наблюдалось. Побочных эффектов обнаружено не было. Гистологические изменений свидетельствовали о снижении эндомизиального фиброза, уменьшении центральной нуклеации, нормализации распределения волокон по размерам и увеличении размера мышц, особенно при высоких дозах вектора [19]. У пациентов наблюдались области с мышечной гипертрофией, но диффузного увеличения четырехглавой мышцы, как в доклинических исследованиях на нечеловекообразных приматах, не было выявлено. Таким образом, примененный подход показал свою перспективность для лечения пациентов с мышечной дистрофией. В испытании NCT02354781 участвовало 3 пациента, которым вводили тот же самый вектор в дозе 1,2×1012 вг/кг/конечность (вг/кг – векторные геномы на килограмм массы тела). Результаты работ еще не опубликованы, так как эксперимент был завершен только в 2020 году.
Известно, что у людей тренировка с отягощением или выносливостью подавляет экспрессию миостатина [7], что позволяет мышцам расти в размерах. Поэтому неудивительно, что подавление функции миостатина считается потенциальным методом для увеличения роста мышц как реакции на тренировку или для стимуляции их роста даже независимо от нагрузки. Именно по этой причине ВАДА ввело средства, модифицирующие функцию миостатина, в Запрещенный список. Из-за свойств, потенциально повышающих эффективность спортсменов, фоллистатин включен в этот список (класс S4, п. 4. «Агенты, предотвращающие активацию рецептора активина IIB», «Гормоны и модуляторы метаболизма») [20]. Несмотря на то, что в настоящее время нет терапевтических средств на основе фоллистатина, тем не менее, была выявлена продажа потенциального контрафактного ингибитора миостатина (Фоллистатин-344) через Интернет [21, 22]. Среди потребителей, такие препараты наиболее популярны у тяжелоатлетов, пауэрлифтеров и бодибилдеров, которые применяют фоллистатин для набора мышечной массы.
Согласно работе [13], большинство продуктов черного рынка представляют собой «фоллистатин-344», меньшее количество маркируется как «фоллистатин-315». Исходя из строения молекулы, теоретически нет никакой разницы между FS315 и FS344 (см. выше). Препарат «FS315» оказался более труднодоступным, и ни один протестированный продукт не содержал фоллистатина вообще. Следовательно, в исследовании были изучены только товары, маркированные как «фоллистатин-344» [13]. Продукты черного рынка покупались у поставщиков, которые в основном продают препараты, связанные с допингом (например, стероиды, пептиды) через Интернет. Как правило, их продукты маркируются как «не для употребления человеком» или «только для исследовательских целей». Тем не менее, исходя из приводимых описаний продуктов и инструкций, очевидно, что субстанции продаются в допинговых целях. Всего авторами было получено 17 различных препаратов «фоллистатин-344» от производителей или дистрибьюторов в Болгарии, Канаде, Китае, Индии, Словакии, Швейцарии, Нидерландах, Великобритании и США. Как оказалось, только 9 из них фактически содержали FS344. Кроме того, авторами [13] был разработан чувствительный аналитический метод, позволяющий, в зависимости от примененного исследуемого объема, обнаружить пептид FS344 в концентрации ~0,1 нг/мл в 10 мл мочи и ~5 нг/мл в 100 мкл сыворотки. Предполагая, что олигомеры FS344 обладают более длительным периодом полувыведения из сыворотки, чем мономеры, сыворотка должна быть предпочтительной аналитической матрицей в антидопинговом контроле злоупотребления FS344. Очевидно, что эти разработки найдут скорейшее применение в тестировании спортсменов на допинг.
Ингибиторы миостатина являются весьма перспективными кандидатами для терапевтического применения при лечении различных нарушений, связанных с нарушением мышечной массы, таких как дистрофия и саркопения. Тем не менее, следует отметить, что фармакологическое вмешательство с целью улучшения спортивных результатов считается неэтичным и противоречит спортивному духу честной и чистой конкуренции. Терапевтическое применение фоллистатина для подавления функции миостатина находится в стадии клинических исследований, предварительные результаты которых выглядят многообещающе.
1. Prado, C.M., Purcell, S.A., Alish, C., Pereira, S.L., Deutz, N.E., Heyland, D.K., Goodpaster, B.H., Tappenden, K.A., Heymsfield, S.B., Implications of low muscle mass across the continuum of care: a narrative review. Annals of Medicine, 2018. 50(8): p. 675-693. DOI: 10.1080/07853890.2018.1511918.
2. McFarlane, C., Sharma, M., Kambadur, R., Role of Myostatin in Skeletal Muscle Growth and Development: Implications for Sarcopenia, in Sarcopenia – Age-Related Muscle Wasting and Weakness: Mechanisms and Treatments, G.S. Lynch, Editor. 2011, Springer Netherlands: Dordrecht. p. 419-447.
3. Owusu-Apenten, R., Bioactive peptides. Applications for improving nutrition and health. 2010, Boca Raton, FL CRC Press, Taylor & Francis Group. p. 416.
4. Mottram, D.R., Mottram, D.R., Chester, N., Drugs in sport. 2018: Routledge.
5. McPherron, A.C., Lawler, A.M., Lee, S.-J., Regulation of skeletal muscle mass in mice by a new TGF-p superfamily member. Nature, 1997. 387(6628): p. 83-90. DOI: 10.1038/387083a0.
6. Schuelke, M., Wagner, K.R., Stolz, L.E., Hübner, C., Riebel, T., Kömen, W., Braun, T., Tobin, J.F., Lee, S.J., Myostatin mutation associated with gross muscle hypertrophy in a child. N Engl J Med, 2004. 350(26): p. 2682-2688. DOI: 10.1056/NEJMoa040933.
7. Matsakas, A., Diel, P., The growth factor myostatin, a key regulator in skeletal muscle growth and homeostasis. International journal of sports medicine, 2005. 26(2): p. 83-89. DOI: 10.1055/s-2004-830451.
8. Rodino-Klapac, L.R., Haidet, A.M., Kota, J., Handy, C., Kaspar, B.K., Mendell, J.R., Inhibition of myostatin with emphasis on follistatin as a therapy for muscle disease. Muscle & Nerve, 2009. 39(3): p. 283-296. DOI: 10.1002/mus.21244.
9. Vassilakos, G., Barton, E.R., Insulin-Like Growth Factor I Regulation and Its Actions in Skeletal Muscle. Comprehensive Physiology, 2019. 9: p. 413-438. DOI: 10.1002/cphy.c180010.
10. Sewald, N., Jakubke, H.-D., Peptides from A to Z. A Concise Encyclopedia. 2008, Darmstadt: WILEY-VCH Verlag GmbH & Co. KGaA. p. 400.
11. Kaneko, H., Subchapter 33C — Follistatin, in Handbook of Hormones, Y. Takei, Ando, H., Tsutsui, K., Editors. 2016, Academic Press: San Diego. p. 298-e233C-292.
12. Identifier: P19883 (FST_HUMAN): the UniProt Knowledgebase (UniProtKB). Available from: https://www.uniprot.org/uniprot/P19883.
13. Reichel, C., Gmeiner, G., Thevis, M., Detection of black market follistatin 344. Drug Testing and Analysis, 2019. 11(11-12): p. 1675-1697. DOI: 10.1002/dta.2741.
14. Inouye, S., Guo, Y., Depaolo, L., Shimonaka, M., Ling, N., Shimasaki, S., Recombinant Expression of Human Follistatin with 315 and 288 Amino Acids: Chemical and Biological Comparison with Native Porcine Follistatin*. Endocrinology, 1991. 129(2): p. 815-822. DOI: 10.1210/endo-129-2-815.
15. US 20070135336 A1, 2007.
16. Mendell, J.R., Translational Medicine: Evolution, Fulfillment, and Belief in Gene Therapy. Human Gene Therapy, 2020. 31(1-2): p. 14-19. DOI: 10.1089/hum.2019.29103.jrm.
17. Haidet, A.M., Rizo, L., Handy, C., Umapathi, P., Eagle, A., Shilling, C., Boue, D., Martin, P.T., Sahenk, Z., Mendell, J.R., Kaspar, B.K., Long-term enhancement of skeletal muscle mass and strength by single gene administration of myostatin inhibitors. Proceedings of the National Academy of Sciences, 2008. 105(11): p. 4318. DOI: 10.1073/pnas.0709144105.
18. Kota, J., Handy, C.R., Haidet, A.M., Montgomery, C.L., Eagle, A., Rodino-Klapac, L.R., Tucker, D., Shilling, C.J., Therlfall, W.R., Walker, C.M., Weisbrode, S.E., Janssen, P.M.L., Clark, K.R., Sahenk, Z., Mendell, J.R., Kaspar, B.K., Follistatin gene delivery enhances muscle growth and strength in nonhuman primates. Sci Transl Med, 2009. 1(6): p. 6ra15-16ra15. DOI: 10.1126/scitranslmed.3000112.
19. Mendell, J.R., Sahenk, Z., Malik, V., Gomez, A.M., Flanigan, K.M., Lowes, L.P., Alfano, L.N., Berry, K., Meadows, E., Lewis, S., Braun, L., Shontz, K., Rouhana, M., Clark, K.R., Rosales, X.Q., Al-Zaidy, S., Govoni, A., Rodino-Klapac, L.R., Hogan, M.J., Kaspar, B.K., A phase 1/2a follistatin gene therapy trial for Becker muscular dystrophy. Molecular Therapy, 2015. 23(1): p. 192-201. DOI: 10.1038/mt.2014.200.
20. Международный Стандарт: Кодекс, Всемирный Антидопинговый. Приложение 1. Запрещенный список. 2020; Available from: https://rusada.ru/upload/iblock/06d/%D0%97%D0%B0%D0%BF%D1%80%D0%B5%D1%89%D0%B5%D0%BD%D0%BD%D1%8B%D0%B9%20%D1%81%D0%BF%D0%B8%D1%81%D0%BE%D0%BA%202020.pdf.
21. Graham, M., Davies, B., Grace, F., Baker, J.J.J.S.M.D.S., Exercise, science and designer doping: Traditional and emerging trends. 2012. 2(113): p. 2161-0673.100011.
22. Зверева, И., Семенистая, Е., Кротов, Г., Родченков, Г., Идентификация допинговых соединений пептидной природы, распространяемых через интернет. Аналитика, 2014(3): p. 58-70.




{kind=link}
{kind=link}
{kind=link}
{kind=link}