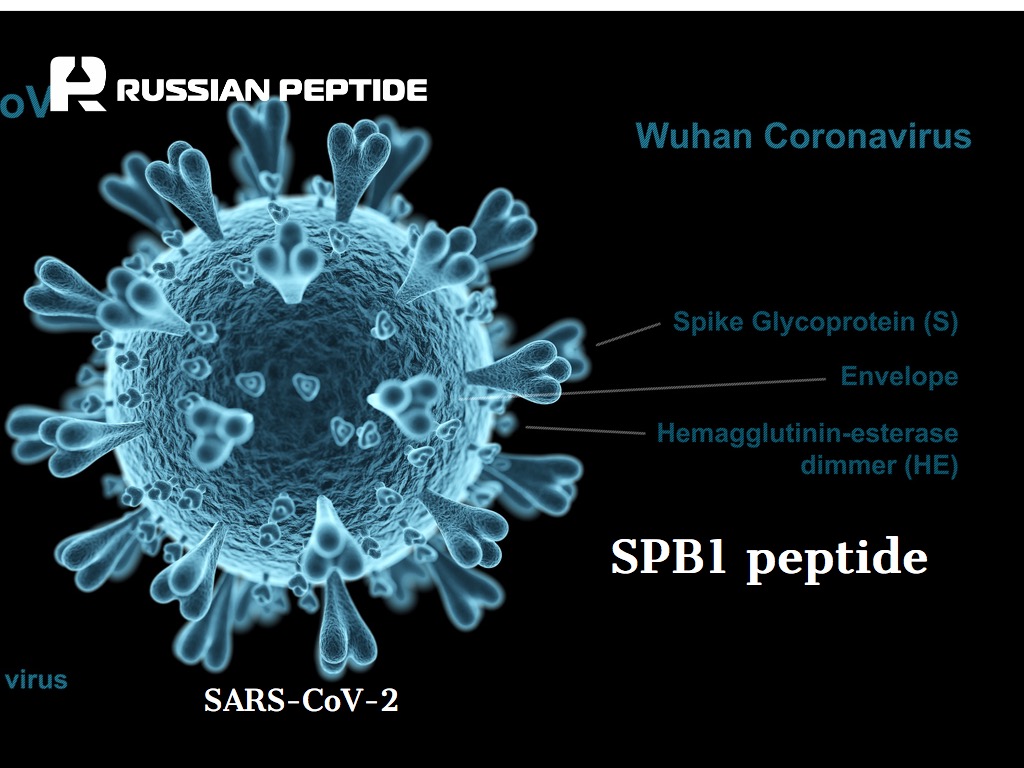
Пептид SPB1 против COVID-19
25.09.2020
Эндогенный антиоксидант Глутатион
06.10.2020
Природный эндогенный микропептид гуманин (Humanin) и его производные защищают клетки от повреждений при нейродегенеративных заболеваниях, включая болезнь Альцгеймера. Пептид также активен при ишемических поражениях сердца, атеросклеротических бляшках, болезнях глаз и в улучшении роста костной ткани. В данном обзоре рассмотрены результаты исследований и представлена история открытия гуманина.
Гуманин, общие сведения и история открытия
Гуманин (Humanin, HN, CAS 330936-69-1) представляет собой природный микропептид с молекулярной массой 2687,27 Да, кодируемый в геномах млекопитающих и включающий 24 остатка: Met-Ala-Pro-Arg-Gly-Phe-Ser-Cys-Leu-Leu-Leu-Leu-Thr-Ser-Glu-Ile-Asp-Leu-Pro-Val-Lys-Arg-Arg-Ala (MAPRGFSCLLLLTSEIDLPVKRRA) (Рис. 1) [1]. Первоначально гуманин был идентифицирован как эндогенный пептид, который защищает нервные клетки от апоптоза (гибели клеток), индуцированного мутантными генами при болезни Альцгеймера. Также сообщалось, что гуманин предотвращает апоптоз клеток PC12 крысы, вызванный снижением уровня сыворотки, и ингибирует апоптоз, препятствуя активации Bcl-2-ассоциированного X-белка (Bax). Было показано, что гуманин может повышать уровень АТФ в лимфоцитах человека; также высказано предположение, что экспрессия гуманина может быть индуцирована в ответ на энергетический кризис в пораженных сосудах связок волокон в мышцах при MELAS синдроме (Mitochondrial Encephalomyopathy, Lactic Acidosis, and Stroke-Like Episodes — митохондриальная энцефаломиопатия с лактат-ацидозом и инсультоподобным синдромом), следовательно, пептид потенциально является терапевтическим кандидатом для лечения MELAS [2, 3]. Гуманин подавляет гибель нейронных клеток, вызванную мутантным белком-предшественником амилоида; подавляет гибель нейрональных клеток, вызванную тремя различными типами генов FAD и β-амилоидом.
Важно отметить, что длина пептида зависит от того, где он секретируется. Если гуманин вырабатывается внутри митохондрий, то будет состоять из 21 аминокислоты [4]. Если же он вырабатывается вне митохондрий, в цитозоле, то гуманин будет включать 24 аминокислотных остатка. Было показано, что оба пептида обладают биологической активностью; всего для человека описано десять близкородственных пептидов [2, 4]. Небольшие гуманиноподобные пептиды представляют собой группу пептидов, обнаруженных в митохондриальной 16S рРНК, которые также обладают ретроградными сигнальными функциями. Экспрессируемый пептид содержит трехвитковую α-спираль и не имеет симметрии [5]. Мутационный анализ гуманина позволил выяснить значимость каждой аминокислоты в его составе (Рис. 1) [4]. Аминокислоты отмечены согласно их функциям, как указано в легенде к рисунку.

Рисунок 1 – Функции определенных аминокислотных остатков в гуманине
Лаборатория Кафедры фармакологии и неврологии Медицинского факультета Университета Кейо (Keio University, Токио, Япония) обнаружила гуманин (MTRNR2) при поиске факторов выживания в непораженном участке мозга пациента с болезнью Альцгеймера [6]. Авторы при помощи библиотеки кДНК проводили поиск генов, которые могли бы противостоять апоптозу, индуцированному мутантной формой предшественника амилоида (APP V642I), который представляет собой ген, мутирующий при одной из форм семейной болезни Альцгеймера. Они также показали, что гуманин может защищать от других семейных генов болезни Альцгеймера, но не может защищать от модели болезни Хантингтона или бокового амиотрофического склероза [6, 7]. Это свидетельствует о том, что гуманин обеспечивает специфическую защиту в моделях болезни Альцгеймера, но не защищает клетки от всех апоптотических событий. Гуманин также взаимодействует с инсулиноподобным белком связывающим фактор роста (IGFBP3), где он может блокировать индуцированный IGFBP3 апоптоз в линиях глиальных клеток, но не в линиях нейрональных клеток или первичных нейронах [8]. Хотя гуманин может блокировать апоптотические эффекты IGFBP3 в глиальных клетках, неожиданно IGFBP3 и гуманин оказывают положительный синергетический эффект на защиту нейронов от апоптоза, индуцированного β-амилоидом [8]. Открытие гуманина интересно многим причины. Это небольшой секретируемый пептид состоящий из 24 или 21 аминокислоты, в зависимости от цитоплазматической или митохондриальной трансляции, соответственно, обладает биологической активностью, хотя и не имеет сигнального пептида для секреции, но может сам действовать как сигнальный пептид [9]. Более интересным является тот факт, что это был первый новый пептид, обнаруженный в митохондриальном геноме с момента его полного секвенирования в 1981 году [4]. Сам митохондриальный геном чрезвычайно мал и компактен (16569 нуклеотидов у человека), без известных интронов и малым количеством некодирующих нуклеотидов между соседними генами; традиционно было известно, что он кодирует только 2 рРНК, 13 мРНК и 22 тРНК. Гуманину соответствует открытая рамка считывания (ORF, последовательность мРНК, не содержащая терминирующих кодонов, которая потенциально может быть транслирована в полипептидную цепь), обнаруженная в гене 16s рРНК. Подобно матрешке, гуманин – это ген в гене внутри генома органеллы внутри клетки; исследователи отнесли его к первому пептиду нового класса митохондриальных пептидов. Гуманин был независимо обнаружен тремя разными лабораториями. Помимо первой опубликованной в 2001 году работы японских исследователей [6], лаборатория Института Бернхема (The Burnham Institute, Калифорния, США) обнаружила гуманин при скрининге белков, которые могут взаимодействовать с Bcl-2-связанным X-белком (Bax), основным белком, участвующим в апоптозе [2]. Международная группа ученых независимо обнаружила гуманин при скрининге белков, которые взаимодействуют с IGFBP3 [8].
Механизм действия гуманина изучен не досконально, однако были опубликованы результаты исследования, в котором продемонстрированы внеклеточные взаимодействия с трехкомпонентным рецептором, состоящим из gp130, WSX1 и CNTFR, а также с рецептором формилпептида 2 (FPR2 из семейства рецепторов, сопряжённых с G-белком, GPCRs) [10, 11]. Внутриклеточное взаимодействие с tBID, BAX, TRIM11 и IGFBP3 также может имеет значение для действия гуманина. [2, 8, 12]. К настоящему времени разработан эффективный метод с использованием твердофазного синтеза для получения гуманина с высоким выходом [13].
Нейропротекторные свойства гуманина
Эксперименты с использованием культивированных клеток продемонстрировали, что гуманин обладает как нейропротекторным, так и цитопротекторным действием, а эксперименты на грызунах показали, что он оказывает защитное действие на моделях болезни Альцгеймера, болезни Хантингтона и инсульта [4, 14].
Исследования на клетках и грызунах показали, что введение гуманина или производных гуманина увеличивает выживаемость и/или улучшает физиологические параметры в моделях болезни Альцгеймера [7, 15].
Как было показано на клетках F11, гибридных клетках первично культивированного нейрона эмбриона крысы (E13) и нейробластомы мыши, гуманин противодействует нейротоксичности различных типов генов семейной болезни Альцгеймера [V642I и K595N/M596L (NL) мутанты белка-предшественника амилоида (APP), M146L-пресенилин (PS)1 и N141I-PS2] и Aβ1–43 с четкой специфичностью действия, неэффективной в отношении нейротоксичности, обусловленной мутациями полиглутаминового повтора Q79 или супероксиддисмутазы 1 [7]. Гуманин может также подавлять нейротоксичность при других поражений, связанных с болезнью Альцгеймера: семейные гены A617G-APP, L648P-APP, A246E-PS1, L286V-PS1, C410Y-PS1 и H163R-PS1, стимуляция APP с помощью антитела против АРР и другие пептиды Aβ (Aβ1–42 и Aβ25–35). При введении в культуральную среду 10 нМ гуманина гибель клеток уменьшалась на ~60% при воздействиях, против которых работал данный пептид (Aβ1–42 и Aβ25–35). На специфичность действия дополнительно указывало, что гуманин не может подавлять нейротоксичность глутаматного или прионного фрагментов. Основные эффекты гуманина против повреждений, связанных с болезнью Альцгеймера, обусловлены наличием Cys8 и Ser14 и доменом от Pro3 до Pro19 [7]. Пептиды, производные гуманина, важны для изучения патофизиологии болезни Альцгеймера и могут открыть новые возможности для разработки терапии при различных формах болезни Альцгеймера.
Для оценки эффективен ли гуманин in vivo в модельных системах болезни Альцгеймера, авторы исследовали in vitro на амнезию, вызванную Aβ25–35 у мышей, влияние HNG, производного гуманина [15].
HNG – мутантный вариант гуманина (S14G-humanin, humanin G, S14G-HN), в котором серин в положении 14 заменен на глицин, т. е, его аминокислотная последовательность следующая: Met-Ala-Pro-Arg-Gly-Phe-Ser-Cys-Leu-Leu-Leu-Leu-Thr-Gly-Glu-Ile-Asp-Leu-Pro-Val-Lys-Arg-Arg-Ala (MAPRGFSCLLLLTGEIDLPVKRRA). Тест в Y-лабиринте показал, что по меньшей мере 50 пмоль HNG при интрацеребровентрикулярной инъекции предотвращают индуцированное нарушение кратковременной и пространственной рабочей памяти; однако 5 нмоль S14A-HN не предотвращал индуцированную амнезию. Эти результаты согласуются с корреляцией структура-функция, показанной ранее in vitro. В задаче по поиску воды HNG предотвращал удлинение латентного периода (времени для поиска воды), наблюдаемого у мышей с амнезией, указывая на то, что HNG также блокирует индуцированное нарушение латентного обучения. Иммуногистохимический анализ показал, что HNG восстанавливает количество холинергических нейронов в базальном переднем мозге и полосатом теле почти до нормального уровня. Кроме того, генистеин, специфический ингибитор тирозинкиназ, блокировал восстановление с помощью HNG после скополаминовой амнезии, что позволяет предположить, что определенные тирозинкиназы участвуют в ингибирующей функции HNG in vivo. Обобщая эти данные, авторы заключили, что HNG обладает протекторной активностью против ухудшения памяти, вызванного поражениями, связанными с БА, in vivo путем активации внутриклеточных нейропротекторных механизмов. Действие HNG in vitro оказалось в 1000 раз сильнее, чем для нативного гуманина.
Гуманин проявляет нейропротективное действие не только по отношению к болезни Альцгеймера, но и прионному поражению, и инсульту [16-18].
В работе [17] авторы обнаружили, что инкубация кортикальных нейронов крысы с 10 мкM гуманина практически полностью предотвращала гибель клеток, индуцированную растворимым фрагментом PrP(118–135), одновременно с ингибированием апоптотических событий. Также изучен мутант гуманина HNG. HNG проявлял 500-кратное увеличение защитной активности в корковых нейронах, тогда как вариант гуманин не проявлял защитного эффекта. Эффекты гуманина и HNG не требовали предварительной инкубации с фрагментом PrP(118–135), что убедительно свидетельствует о том, что эти пептиды защищают клетки независимо от прямого взаимодействия с прионным пептидом. Напротив, гуманин не влиял на гибель клеток, индуцированную фибриллярным пептидом PrP(106–126).
Гуманин в концентрации 1–10 мкМ многократно ослаблял апоптоз клеток линии PC12, вызванный белками DRPLA с расширенными полиглутаминовыми повторами [16].
В исследовании [18] изучался нейрозащитный эффект HNG (производного гуманина) на очаговую ишемию головного мозга/реперфузионное повреждение у мышей. Мышам производили окклюзию средней мозговой артерии в течение 75 минут с последующей 24-часовой реперфузией, предварительно вводя 0,1 мкг HNG (внутрицеребровентрикулярно) за 30 минут до ишемии или с постобработкой через 0, 2, 4 и 6 часов после ишемии; или предварительно вводя 1 мкг HNG (внутрибрюшинно) за 1 час до ишемии. Предварительное применение 0,1 мкг HNG (внутрицеребровентрикулярно) за 30 минут до ишемии уменьшала объем церебрального инфаркта с 56,2% до 26%. Постобработка через 4 часа реперфузии снизила объем церебрального инфаркта до 45,6%. Предварительная обработка 1 мкг HNG (внутрибрюшинно) за 1 час до ишемии или после лечения через 2 часа реперфузии значительно уменьшала объем инфаркта мозга. HNG также существенно улучшил неврологическую функцию и ингибировал апоптоз нейронов, а также активацию полимеразы поли(АДФ-рибозы). Значительное снижение фосфо-ERK наблюдалось у мышей, получавших HNG, тогда как уровни фосфо-JNK и фосфо-p38 не изменялись. Результаты демонстрируют, что HNG защищает от церебральной ишемии/реперфузионного повреждения у мышей. HNG обеспечивает нейрозащиту in vivo, по крайней мере частично, путем ингибирования активации ERK, что предполагает потенциальную терапевтическую роль HNG в лечении инсульта [18].
Гуманин при нервно-мышечных заболеваниях
Гуманин первоначально был идентифицирован как эндогенный пептид, который защищает нейрональные клетки от апоптоза, вызванного различными типами поражений, связанных с болезнью Альцгеймера. Исследователи предположили, что этот пептид может защищать клетки с дефицитом энергии при митохондриальных нарушениях[3]. Было обнаружено увеличение экспрессии гуманина в скелетных мышцах у пациентов с митохондриальной энцефаломиопатией с лактоацидозом и эпизодами инсульта (MELAS). Гуманин обнаруживался во всех рваных красных волокнах (RRF, гистологический признак многих нервно-мышечных заболеваний) и некоторых других, причем большинство из них были волокнами типа 1, обычно требующими большей энергии, чем волокна типа 2. Пептид в этих волокнах был локализован в митохондриях. Экспрессия гуманина также увеличивалась в мелких артериях. Эксперименты на мышечных клетках TE671 показали возможность того, что синтетический гуманин увеличивает клеточные уровни АТФ, напрямую воздействуя на митохондрии. На основании этих результатов авторы предположили, что экспрессия пептида может быть индуцирована в ответ на энергетический кризис в пораженных волокнах и сосудах в мышцах при синдроме MELAS и его введение может стать путем к разработке терапии этого заболевания [3].
Кроме указанных нейропротекторных свойств, гуманин способен предохранять от окислительного стресса, формирования атеросклеротических бляшек и ишемии [18-20]. Описаны полезные метаболические эффекты – гуманин может повышать выживаемость β-клеток поджелудочной железы, что полезно при диабете типа 1[21], и улучшает восприимчивость к инсулину, что благоприятно действует в случае диабета типа 2[22].
Гуманин при инфаркте миокарда
В работе [20] оценили эффекты HNG, мощного аналога гуманина, на мышиной модели ишемии и реперфузии миокарда in vivo. HNG внутрибрюшинно за 1 час до или во время реперфузии. Распространенность инфаркта миокарда в каждой зоне риска оценивали через 24 часа. Функцию левого желудочка оценивали через 1 неделю после ишемии с помощью 2D-эхокардиографии высокого разрешения. HNG уменьшал размер инфаркта относительно зоны риска в зависимости от дозы, с максимальным уменьшением при дозе 2 мг/кг. Применение HNG увеличило фракцию выброса левого желудочка и сохранила постишемические размеры левого желудочка (конечный диастолический и конечный систолический), что привело к улучшению сердечной функции. Лечение HNG значительно увеличивало фосфорилирование AMPK и фосфорилирование эндотелиальной синтазы оксида азота в сердце и снижало уровни Bcl-2-ассоциированного X-белка и B-клеточной лимфомы-2 после ишемии и реперфузии миокарда. HNG улучшал выживаемость кардиомиоцитов и снижал апоптоз в ответ на даунорубицин in vitro. Эти данные показывают, что HNG обеспечивает кардиопротекцию на мышиной модели ишемии и реперфузии миокарда, потенциально за счет активации AMPK-эндотелиальной синтазы оксида азота, опосредованной синтазой передачи сигналов и регуляции апоптотических факторов. HNG может представлять собой новое средство для лечения острого инфаркта миокарда.
На людях гуманин еще не исследовали, но в 2020 году завершилось клиническое исследование NCT03431844, цель которого было определить концентрацию гуманиноподобного пептида в ткани миокарда и в крови, а также выяснить, связан ли уровень его концентрации с ранним возникновением и частотой осложнений после операции на сердце. К настоящему моменту результаты работы еще не опубликованы.
Гуманин и диабет
В экспериментах на крысах аналог гуманина способен нормализовать концентрацию глюкозы и снижает симптомы диабета. Ухудшение действия инсулина является метаболическим признаком старения и участвует в развитии возрастных заболеваний, включая сахарный диабет типа 2 (T2DM) и болезнь Альцгеймера. Как показали исследователи в работе [22], гуманин в дозе 20 мкг влияет на чувствительность к инсулину. Используя зажимную технологию, авторы исследовали роль центрального и периферического гуманина на действие инсулина в крысиной модели. Непрерывная инфузия значительно улучшала общую чувствительность к инсулину. Центральные эффекты гуманина на действие инсулина были связаны с активацией гипоталамической передачи сигналов STAT-3; эффекты, были сведены на нет совместным ингибированием гипоталамического STAT-3. Периферические внутривенные инфузии новых и сильнодействующих производных гуманина воспроизводили инсулино-сенсибилизирующие эффекты центрального введения пептида. Однократное применение сильнодействующим аналогом F6AHN, который имеет единственную аминокислотную замену аланина на фенилаланин в положении 6, что предотвращает связывание IGFBP3, значительно, практически в два раза, снижало уровень глюкозы в крови у крыс с диабетом Цукера и ожирением. Основываясь на связи гуманина с возрастными заболеваниями, авторы выявили, что количество обнаруживаемого гуманина в гипоталамусе, скелетных мышцах и коре головного мозга снижалось с возрастом у грызунов, а уровни циркулирующего пептида уменьшались с возрастом у людей и мышей. Авторы пришли к выводу, что снижение концентрации гуманина может играть роль в патогенезе возрастных заболеваний, включая болезнь Альцгеймера и диабетом типа 2.
Гуманин подавляет апоптоз β-клеток на 60%, а также задерживает и облегчает диабет у мышей, не страдающих ожирением при 6-недельной терапии внутрибрюшинными инъекциями гуманина по 0,7 мг/кг/день [21].
Влияние гуманина на эпителий сетчатки
Возрастная дегенерация желтого пятна (AMD) является основной причиной тяжелой и необратимой потери зрения и характеризуется прогрессирующей дегенерацией сетчатки, приводящей к потере центрального зрения. Пигментный эпителий сетчатки является критическим местом патологии AMD. Митохондрии и эндоплазматический ретикулум, которые находятся в анатомической близости друг к другу, являются мишенями окислительного стресса и стресса эндоплазматического ретикулума (ER), соответственно, и способствуют прогрессированию AMD. В работе [23] представлены доказательства перекрестного взаимодействия ER-митохондрий в пигментном эпителии сетчатки при стрессе эндоплазматического ретикулума и сигнальных путей апоптотической гибели клеток. Авторами обсуждается роль гуманина в модуляции ER и окислительного стресса в пигментном эпителии сетчатки. Гуманин защищал клетки эндоплазматического ретикулума от окислительной и вызванной стрессом эндоплазматического ретикулума гибели клеток посредством активации митохондриального GSH, ингибирования генерации активных форм кислорода и активации каспаз 3 и 4. Авторами осуждаются лежащие в основе механизмы ER-митохондриального перекрестного взаимодействия и модуляции экзогенным гуманином. Терапевтическое использование гуманина и родственных пептидов потенциально может оказаться ценным подходом к лечению AMD.
Позже было обнаружено, что наночастицы гуманина являются эффективным средством для снижения патологических факторов, характерных для возрастной дегенерации желтого пятна [24]. AGA-HNG был синтезирован и инкапсулирован в наночастицы хитозана с радиусом 346 нм, которые затем были охарактеризованы по их размеру, эффективности инкапсуляции (и высвобождению лекарства. Их способность подавлять секрецию VEGF и защищать от окислительного апоптоза изучалась in vitro с использованием клеток пигментного эпителия сетчатки человека ARPE-19. Наночастицы проявляли такие же свойства против VEGF (фактор роста эндотелия сосудов) и окислительную защиту, как и свободный пептид, при этом проявляя превосходные фармацевтические характеристики, включая биосовместимость и скорость высвобождения лекарства. AGA-HNG показал большие перспективы в опосредовании апоптоза в гипоксических клетках, выживаемость была на 50% выше, чем при применении HNG, особенно в высокой дозе 10 мкМ. Клетки, обработанные AGA-HNG, показали в среднем на 232% большую жизнеспособность, чем контроль, независимо от концентрации или режима обработки. Также значительно снизилась экспрессия VEGF in vitro со сниженной цитотоксичностью по сравнению со свободным лекарством [24]. Способность этой системы доставки лекарств уменьшать апоптоз сетчатки с желаемыми фармакокинетическими и биосовместимыми свойствами делает ее многообещающим вариантом лечения AMD.
Влияние гуманина на нарушения роста костей
В исследовании [25] влияние глюкокортикоидов и гуманина на рост костей оценивалось на нескольких различных экспериментальных моделях. Авторы обнаружили, что синтетический аналог гуманина, HNG, полностью избавил от нарушения роста костей, вызванного дексаметазоном, и что мыши со сверхэкспрессией гуманина (HNtg) были устойчивы к нарушению роста, вызванному глюкокортикоидами. Кроме того, HNG помогает избавиться от индуцированного глюкокортикоидами апоптоза в пластинке роста на модели хронического воспаления. HNG в концентрации 100 нМ подавлял апоптоз как в пластинке роста, так и в суставном хряще, рост кости увеличивался на 35% по сравнению с обработанной только дексаметазоном. Важно отметить, что HNG не влияет на противовоспалительный эффект дексаметазона. Обработка дексаметазоном мышей дикого типа сопровождалась подавлением общей высоты пластинки роста и уменьшением высоты зоны покоя и пролиферации. HNG смог восстановить эту потерю высоты ростовой пластинки у мышей дикого типа. У мышей HNtg обработка дексаметазоном не влияла на высоту пластинки роста. На основании этих результатов авторы предположили, что гуманин может защищать от апоптоза в пластинке роста. Они также обнаружили, что экспрессия Ihh, важная для пролиферации и дифференцировки хондроцитов, подавлялась в ростовых пластинах мышей дикого типа, и что HNG был способен восстанавливать уровни Ihh в ростовых пластинах.
Антинекротические эффекты гуманина
Запатентован способ лечения пациентов, страдающих заболеваниями, характеризующимися некрозом тканей, путем введения пациенту ингибитора некроза и/или гуманина в дозировке 50–150 мг/кг массы тела/день [26]. Авторы проверили действие пептида и его производных in vitro на клеточных линиях U937, PC12, NSC348 и HL1 при поражении их раствором цианида калия, стауроспорином и олигомицином. Выживаемость клеток, измеренная по концентрации лактатдегидрогеназы (стабильный цитозольный фермент из лизированных клеток), возрастала на 20–30% под действием пептидов.
Связь гуманина с онкогенезом
Антиапоптотические свойства гуманина вызывало у исследователей озабоченность по поводу того, способствует ли он развитию рака и помогает ли выживанию раковых клеток. Основываясь том, что обнаружено повышение экспрессии митохондриального гена 16S рРНК в неходжкинской лимфоме, что потенциально увеличивает экспрессию гуманина, было предположено, что гуманин является онкопептидом [27]. Кроме того, гуманин был иммунологически обнаружен в сыворотке некоторых пациентов с кожной Т-клеточной лимфомой, но не у здоровых субъектов [28]. Используя дрожжевую модель, группа ученых определила MPP8, который представляет собой известный онкобелок, играющий роль в подвижности опухоли и инвазии, в качестве партнера по связыванию гуманина [29], таким образом, указывая на потенциальную роль гуманина в онкогенезе. Исследование показало сверхэкспрессию гуманина и его изоформ в химиорезистентных опухолевых тканях желудка пациентов. Используя qRT-PCR, авторы подтвердили повышенные уровни мРНК генов HN1, HN3, HN6 и HN10, предполагая потенциальную роль гуманина в развитии химиорезистентности в опухолевых клетках желудка. Ограничением исследования, как отметили авторы, было отсутствие измерений уровня белка, которые невозможно было провести из-за отсутствия антител, специфичных для изоформ.
Кроме того, было показано, что гуманин сверхэкспрессируется в опухолевых клетках мочевого пузыря, и, поскольку пептид присутствует в крови, было высказано предположение, что циркулирующий гуманин потенциально может служить новым биомаркером в диагностике карциномы [30].
Напротив, было продемонстрировано, что при введении HNG с бортезомибом (противоопухолевое средство, протеасомный ингибитор), HNG предотвращал индуцированное бортезомибом нарушение роста костей у мышей с моделями ксенотрансплантата опухоли человека [31], не влияя на его химиотерапевтические эффекты. В этом эксперименте мыши получали один 2-недельный цикл (2 инъекции в неделю) бортезомиба (0,8 мг/кг или 1,0 мг/кг), или HNG (1 мкг/мышь), или комбинацию HNG/бортезомиб или физраствор. Рост кости увеличился с 0,09–0,11 мм/день в группе бортезомиба до 0,15–0,16 мм/день при введении в комбинации с HNG. Они также показали на ксенотрансплантате опухоли человека и клеточных линиях, что HNG предотвращает индуцированный бортезомибом апоптоз, предотвращая активацию Bax и PARP. Что еще более интересно, один только HNG задерживал рост опухоли и время удвоения опухоли при раковых заболеваниях, таких как медуллобластома и нейробластома in vivo. Авторы предположили, что увеличение Bax в хондроцитах по сравнению с клетками нейробластомы человека может быть основой защиты хондроцитов HNG после применения бортезомиба.
Также сообщалось, что HNG защищает от вызванной циклофосфамидом потери мужских GC и лейкоцитов и усиливает индуцированное циклофосфамидом подавление метастазов рака на модели метастатической меланомы легких у мышей [32]. Поэтому было высказано предположение, что аналоги гуманина могут быть полезными адъювантами к химиотерапии за счет уменьшения побочных эффектов. Фактически, было высказано предположение, что они могут увеличивать ответ опухолевых клеток на бортезомиб и циклофосфамид.
Однако недавние результаты противоречат этим выводам [33]. Ингибирование эндогенного гуманина путем внутриопухолевой инъекции бакуловирусных генотерапевтических векторов, кодирующих короткую шпилечную РНК, нацеленную на гуманин, усиливало экспрессию Bax, увеличивало скорость апоптоза опухоли, подавляло рост опухоли и увеличивало выживаемость моделей ксенотрансплантата пролактиномы. Кроме того, предварительные результаты показывают, что системное введение гуманина способствует прогрессированию опухоли и химиорезистентности на мышиных моделях рака молочной железы, что ставит под сомнение безопасность системного введения пептида или его аналогов для лечения рака.
Данные о связи гуманина с онкогенезом и метастазированием рака выглядят противоречиво. Очевидно, что требуются дальнейшие доклинические исследования эффектов пептида при онкозаболеваниях.
Заключение
Таким образом, из приведенных выше данных видно, что гуманин и его производные обладают широким спектром цитопротекторных эффектов, что позволяет предположить потенциальную полезность пептида в разработке лекарственных средств для терапии болезни Альцгеймера и других нейродегенеративных заболеваний, диабета, болезней сердца, глаз и других.
1. Sewald, N., Jakubke, H.-D., Peptides from A to Z. A Concise Encyclopedia. 2008, Darmstadt: WILEY-VCH Verlag GmbH & Co. KGaA. p. 400.
2. Guo, B., Zhai, D., Cabezas, E., Welsh, K., Nouraini, S., Satterthwait, A.C., Reed, J.C., Humanin peptide suppresses apoptosis by interfering with Bax activation. Nature, 2003. 423(6938): p. 456-461. DOI: 10.1038/nature01627.
3. Kariya, S., Hirano, M., Furiya, Y., Sugie, K., Ueno, S., Humanin detected in skeletal muscles of MELAS patients: a possible new therapeutic agent. Acta Neuropathologica, 2005. 109(4): p. 367-372. DOI: 10.1007/s00401-004-0965-5.
4. Kelvin, Y., Changhan, L., Hemal, M., Pinchas, C., The emerging role of the mitochondrial-derived peptide humanin in stress resistance. Journal of Molecular Endocrinology, 2013. 50(1): p. R11-R19. DOI: 10.1530/JME-12-0203.
5. Benaki, D., Zikos, C., Evangelou, A., Livaniou, E., Vlassi, M., Mikros, E., Pelecanou, M., Solution structure of humanin, a peptide against Alzheimer’s disease-related neurotoxicity. Biochemical and Biophysical Research Communications, 2005. 329(1): p. 152-160. DOI: https://doi.org/10.1016/j.bbrc.2005.01.100.
6. Hashimoto, Y., Niikura, T., Tajima, H., Yasukawa, T., Sudo, H., Ito, Y., Kita, Y., Kawasumi, M., Kouyama, K., Doyu, M., Sobue, G., Koide, T., Tsuji, S., Lang, J., Kurokawa, K., Nishimoto, I., A rescue factor abolishing neuronal cell death by a wide spectrum of familial Alzheimer’s disease genes and Abeta. Proceedings of the National Academy of Sciences of the United States of America, 2001. 98(11): p. 6336-6341. DOI: 10.1073/pnas.101133498.
7. Hashimoto, Y., Niikura, T., Ito, Y., Sudo, H., Hata, M., Arakawa, E., Abe, Y., Kita, Y., Nishimoto, I., Detailed characterization of neuroprotection by a rescue factor humanin against various Alzheimer’s disease-relevant insults. The Journal of neuroscience : the official journal of the Society for Neuroscience, 2001. 21(23): p. 9235-9245. DOI: 10.1523/JNEUROSCI.21-23-09235.2001.
8. Ikonen, M., Liu, B., Hashimoto, Y., Ma, L., Lee, K.-W., Niikura, T., Nishimoto, I., Cohen, P., Interaction between the Alzheimer’s survival peptide humanin and insulin-like growth factor-binding protein 3 regulates cell survival and apoptosis. Proceedings of the National Academy of Sciences, 2003. 100(22): p. 13042. DOI: 10.1073/pnas.2135111100.
9. Yamagishi, Y., Hashimoto, Y., Niikura, T., Nishimoto, I., Identification of essential amino acids in Humanin, a neuroprotective factor against Alzheimer’s disease-relevant insults. Peptides, 2003. 24(4): p. 585-595. DOI: https://doi.org/10.1016/S0196-9781(03)00106-2.
10. Hashimoto, Y., Kurita, M., Aiso, S., Nishimoto, I., Matsuoka, M., Humanin Inhibits Neuronal Cell Death by Interacting with a Cytokine Receptor Complex or Complexes Involving CNTF Receptor α/WSX-1/gp130. Molecular Biology of the Cell, 2009. 20(12): p. 2864-2873. DOI: 10.1091/mbc.e09-02-0168.
11. Ying, G., Iribarren, P., Zhou, Y., Gong, W., Zhang, N., Yu, Z.-X., Le, Y., Cui, Y., Wang, J.M., Humanin, a Newly Identified Neuroprotective Factor, Uses the G Protein-Coupled Formylpeptide Receptor-Like-1 as a Functional Receptor. The Journal of Immunology, 2004. 172(11): p. 7078. DOI: 10.4049/jimmunol.172.11.7078.
12. Niikura, T., Hashimoto, Y., Tajima, H., Ishizaka, M., Yamagishi, Y., Kawasumi, M., Nawa, M., Terashita, K., Aiso, S., Nishimoto, I., A tripartite motif protein TRIM11 binds and destabilizes Humanin, a neuroprotective peptide against Alzheimer’s disease-relevant insults. European Journal of Neuroscience, 2003. 17(6): p. 1150-1158. DOI: 10.1046/j.1460-9568.2003.02553.x.
13. Zikos, C., Evangelou, A., Benaki, D., Mikros, E., Pelecanou, M., Livaniou, E., Evangelatos, G.P., High-yield solid-phase synthesis of humanin, an Alzheimer’s disease associated 24-mer peptide, and humanin analogues. Collection of Czechoslovak Chemical Communications, 2015. 8: p. 107-111.
14. Kim, S.-J., Xiao, J., Wan, J., Cohen, P., Yen, K., Mitochondrially derived peptides as novel regulators of metabolism. The Journal of Physiology, 2017. 595(21): p. 6613-6621. DOI: 10.1113/JP274472.
15. Tajima, H., Kawasumi, M., Chiba, T., Yamada, M., Yamashita, K., Nawa, M., Kita, Y., Kouyama, K., Aiso, S., Matsuoka, M., Niikura, T., Nishimoto, I., A humanin derivative, S14G-HN, prevents amyloid-β-induced memory impairment in mice. Journal of Neuroscience Research, 2005. 79(5): p. 714-723. DOI: 10.1002/jnr.20391.
16. Kariya, S., Hirano, M., Nagai, Y., Furiya, Y., Fujikake, N., Toda, T., Ueno, S., Humanin attenuates apoptosis induced by DRPLA proteins with expanded polyglutamine stretches. Journal of Molecular Neuroscience, 2005. 25(2): p. 165-169. DOI: 10.1385/JMN:25:2:165.
17. Sponne, I., Fifre, A., Koziel, V., Kriem, B., Oster, T., Pillot, T., Humanin rescues cortical neurons from prion-peptide-induced apoptosis. Molecular and Cellular Neuroscience, 2004. 25(1): p. 95-102. DOI: https://doi.org/10.1016/j.mcn.2003.09.017.
18. Xu, X., Chua Chu, C., Gao, J., Hamdy Ronald, C., Chua Balvin, H.L., Humanin Is a Novel Neuroprotective Agent Against Stroke. Stroke, 2006. 37(10): p. 2613-2619. DOI: 10.1161/01.STR.0000242772.94277.1f.
19. Zacharias, D.G., Kim, S.G., Massat, A.E., Bachar, A.R., Oh, Y.K., Herrmann, J., Rodriguez-Porcel, M., Cohen, P., Lerman, L.O., Lerman, A., Humanin, a Cytoprotective Peptide, Is Expressed in Carotid Artherosclerotic Plaques in Humans. PLOS ONE, 2012. 7(2): p. e31065. DOI: 10.1371/journal.pone.0031065.
20. Muzumdar, R., H., Huffman, D.M., Calvert, J.W., Jha, S., Weinberg, Y., Cui, L., Nemkal, A., Atzmon, G., Klein, L., Gundewar, S., Ji Sang, Y., Lavu, M., Predmore, B.L., Lefer, D.J., Acute Humanin Therapy Attenuates Myocardial Ischemia and Reperfusion Injury in Mice. Arteriosclerosis, Thrombosis, and Vascular Biology, 2010. 30(10): p. 1940-1948. DOI: 10.1161/ATVBAHA.110.205997.
21. Hoang, P.T., Park, P., Cobb, L.J., Paharkova-Vatchkova, V., Hakimi, M., Cohen, P., Lee, K.-W., The neurosurvival factor Humanin inhibits β-cell apoptosis via signal transducer and activator of transcription 3 activation and delays and ameliorates diabetes in nonobese diabetic mice. Metabolism — Clinical and Experimental, 2010. 59(3): p. 343-349. DOI: 10.1016/j.metabol.2009.08.001.
22. Muzumdar, R.H., Huffman, D.M., Atzmon, G., Buettner, C., Cobb, L.J., Fishman, S., Budagov, T., Cui, L., Einstein, F.H., Poduval, A., Hwang, D., Barzilai, N., Cohen, P., Humanin: A Novel Central Regulator of Peripheral Insulin Action. PLOS ONE, 2009. 4(7): p. e6334. DOI: 10.1371/journal.pone.0006334.
23. Minasyan, L., Sreekumar, P.G., Hinton, D.R., Kannan, R., Protective Mechanisms of the Mitochondrial-Derived Peptide Humanin in Oxidative and Endoplasmic Reticulum Stress in RPE Cells. Oxidative Medicine and Cellular Longevity, 2017.: p. 1675230. DOI: 10.1155/2017/1675230.
24. Solanki, A., Smalling, R., Parola, A.H., Nathan, I., Kasher, R., Pathak, Y., Sutariya, V., Humanin Nanoparticles for Reducing Pathological Factors Characteristic of Age-Related Macular Degeneration. Current Drug Delivery, 2019. 16(3): p. 226-232. DOI: 10.2174/1567201815666181031163111.
25. Celvin, B., Prevention of growth failure caused by glucocorticoids and inflammation. Ph.D. 2019 Karolinska Institutet: Stockholm, Sweden. p. 62.
26. WO 2011104708 A2, 2011.
27. Maximov, V., Martynenko, A., Hunsmann, G., Tarantul, V., Mitochondrial 16S rRNA gene encodes a functional peptide, a potential drug for Alzheimer’s disease and target for cancer therapy. Medical Hypotheses, 2002. 59(6): p. 670-673. DOI: https://doi.org/10.1016/S0306-9877(02)00223-2.
28. Hartmann, T.B., Mattern, E., Wiedemann, N., Doorn, R.v., Willemze, R., Niikura, T., Hildenbrand, R., Schadendorf, D., Eichmüller, S.B., Identification of selectively expressed genes and antigens in CTCL. Experimental Dermatology, 2008. 17(4): p. 324-334. DOI: 10.1111/j.1600-0625.2007.00637.x.
29. Kokura, K., Sun, L., Bedford, M.T., Fang, J., Methyl-H3K9-binding protein MPP8 mediates E-cadherin gene silencing and promotes tumour cell motility and invasion. The EMBO Journal, 2010. 29(21): p. 3673-3687. DOI: 10.1038/emboj.2010.239.
30. Omar, N.N., Tash, R.F., Shoukry, Y., ElSaeed, K.O., Breaking the ritual metabolic cycle in order to save acetyl CoA: A potential role for mitochondrial humanin in T2 bladder cancer aggressiveness. Journal of the Egyptian National Cancer Institute, 2017. 29(2): p. 69-76. DOI: https://doi.org/10.1016/j.jnci.2017.04.001.
31. Eriksson, E., Wickström, M., Perup, L.S., Johnsen, J.I., Eksborg, S., Kogner, P., Sävendahl, L., Protective Role of Humanin on Bortezomib-Induced Bone Growth Impairment in Anticancer Treatment. JNCI: Journal of the National Cancer Institute, 2014. 106(3). DOI: 10.1093/jnci/djt459.
32. Jia, Y., Ohanyan, A., Lue, Y.-H., Swerdloff, R.S., Liu, P.Y., Cohen, P., Wang, C., The effects of humanin and its analogues on male germ cell apoptosis induced by chemotherapeutic drugs. Apoptosis, 2015. 20(4): p. 551-561. DOI: 10.1007/s10495-015-1105-5.
33. Zuccato, C.F., Asad, A.S., Nicola Candia, A.J., Gottardo, M.F., Moreno Ayala, M.A., Theas, M.S., Seilicovich, A., Candolfi, M., Mitochondrial-derived peptide humanin as therapeutic target in cancer and degenerative diseases. Expert Opinion on Therapeutic Targets, 2019. 23(2): p. 117-126. DOI: 10.1080/14728222.2019.1559300.
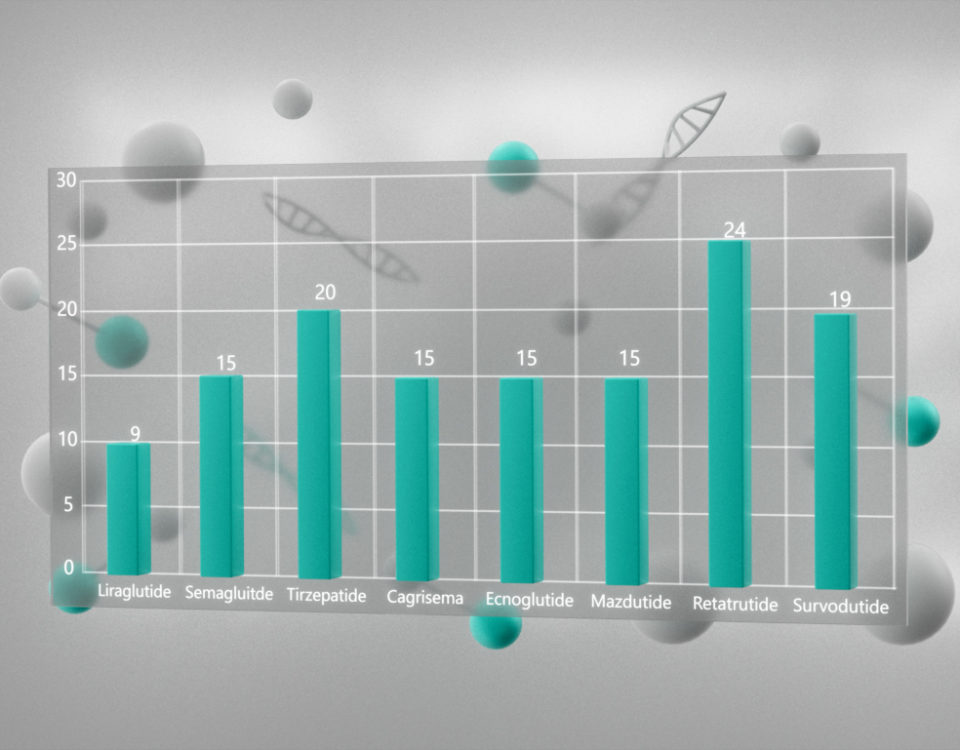




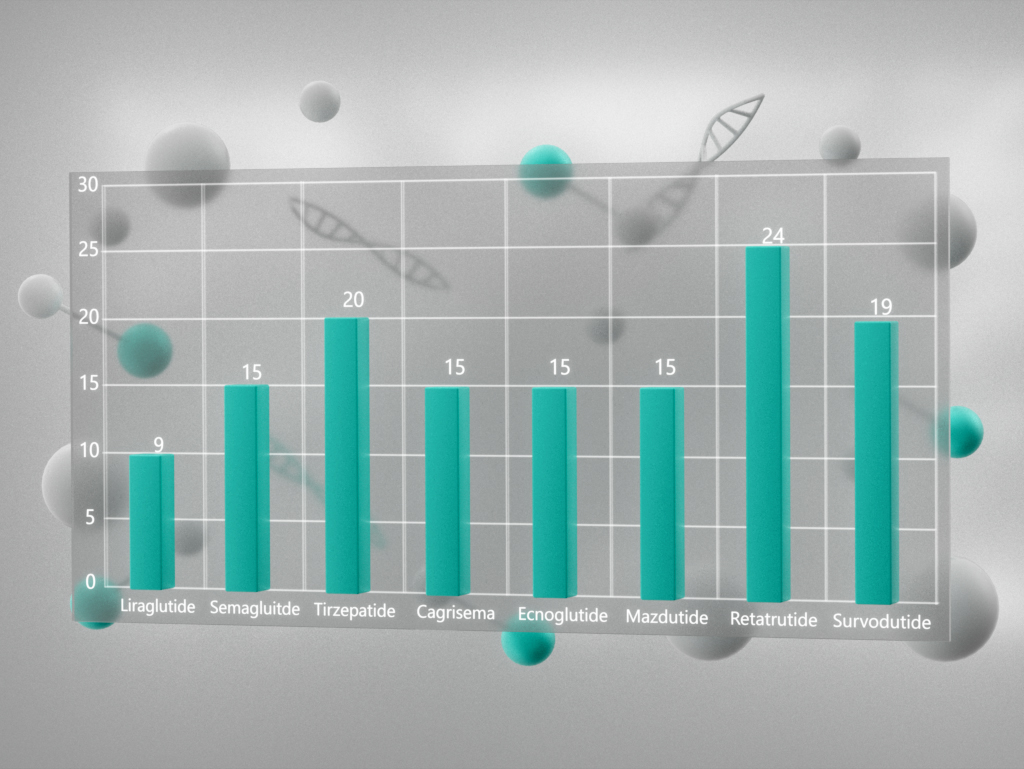{kind=link}
{kind=link}
{kind=link}
{kind=link}