
Пептиды эзрина и HEP-1 против COVID-19
08.06.2020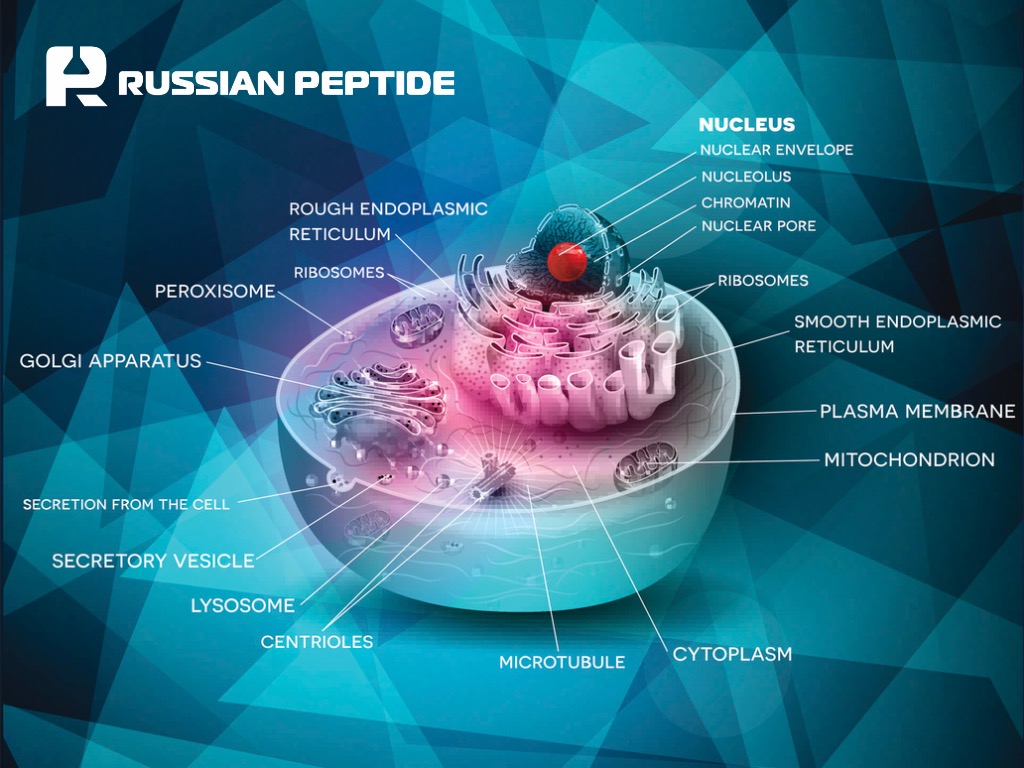
Релизинг-пептиды гормона роста (GHRPs) и цитопротекторные свойства
21.06.2020
Теломеры, длина теломер, теломерный тест, биологический возраст, эпиталон, эпиталамин, мелатонин, тест и норма мелатонина
Теломеры – это концевые фрагменты линейных хромосом. Структурно, теломеры представляют собой повторяющиеся участки некодирующей ДНК, которые служат для защиты хромосом от распознавания клеточным механизмом как поврежденной ДНК, от негомологичного присоединения и пр. У людей теломеры состоят из гексануклеотидного повтора TTAGGG на 3-й цепи (называемого G-обогащенной цепью или G-цепи) и AATCCC на 5-й цепи (называемого C-обогащенной цепью, или С-нить) [1]. Фермент, который поддерживает длину теломер, известен как теломераза. Основная ее функция – катализировать de novo синтез теломер, удлиняя их. Активность теломеразы можно измерить в лабораторных условиях, используя анализ на основе полимеразной цепной реакции (ПЦР), называемый протоколом амплификации теломерных повторов (Telomerase Repeat Amplification Protocol, TRAP). Большой интерес вызывает использование теломеразы в качестве противораковой мишени и маркера раннего выявления рака. Когда теломеры имеют достаточную длину, регуляторные белки и защитные структуры ингибируют ассоциацию теломеразы с концом теломеры. Напротив, когда теломеры короткие, они могут стать доступными для фермента теломеразы из-за потери вышеупомянутых структур или диссоциации теломер-связывающих белков или их регуляторов [1]. Теломеры укорачиваются при репликативном старении клетки, что было показано Хейфликом (LeonardHayflick), подтверждает связь этого явления с общим старением. Клетки человека in vitro имеют конечное значение количества делений (предел Хейфлика). Теломеразная теория имеет в своей основе корреляцию уменьшения пролиферативного потенциала клеток тканей с возрастом человека и критического уменьшения длины теломер при деления клеток; соматические клетки не обладают теломеразной активностью. Результаты анализа длины теломер являются характеристикой биологического возраста, то есть уровень физиологического (функциональность) и морфологического (конституцию) развития человеческого тела, который может не соответствовать фактическому (хронологическому) возрасту как с большую, так и в меньшую сторону.

Длина теломер – важнейший маркер старения, поэтому были созданы специальные тесты для ее определения. Долгое время считалось невозможным измерить теломеры в ДНК позвоночных путем амплификации ПЦР с олигонуклеотидными праймерами, предназначенными для гибридизации с повторами TTAGGG и CCCTAA, однако в 2002 году были разработаны специальные праймеры, реагенты для проведения ПЦР, которые позволили просто и быстро измерять теломеры [2]. Позже метод был усовершенствован так, что стал позволять оценивать длину единичных теломер. Тест получил название STELA (Single Telomere Length Analysis) [3]. В настоящее время известно не менее семи различных вариантов определения длины теломер, каждый из которых имеет свои преимущества и недостатки (подробнее см. обзор [4]). Стали коммерчески доступны специальные киты (наборы реактивов для ПЦР), позволяющие анализировать длину теломер в любой лаборатории, оснащенной соответствующим оборудованием, на котором работает квалифицированный персонал. Цены на такие киты значительно варьируются в зависимости от производителя. Например, «Absolute Human Telomere Length Quantification qPCR Assay Kit» на сто реакций, предлагаемый фирмой ScienCell Research Laboratories стоит 214 USD, а «TeloTAGGG™ Telomere Length Assay», достаточный для проведения 50 анализов, от компании Merck – 1327 USD. Но даже при использовании дорогих реактивов, стоимость проведения одного теста вполне доступна для любого желающего и составляет 10–20 тыс. рублей в различных коммерческих лабораториях.

Некоторые препараты, например, Эпиталон, способны увеличивать длину теломер, что может положительно сказаться на биологическом возрасте человека. Рассмотрим Эпиталон и его положительные эффекты более подробно.
Эпиталон (Epitalon, AE-0 peptide, Epithalon, CAS 307297-39-8) это препарат, представляющий собой синтетический пептид, аминокислотная последовательность которого состоит из четырех остатков – Ala-Glu-Asp-Gly(AEDG). Данный тетрапептид был создан в результате изучения аминокислотного состава эпиталамина в 2002 году [5]. Эпиталамин – биорегулятор нейроэндокринной системы [6, 7] и является фармакопейным лекарственным средством, которое представляет собой вытяжку из шишковидного тела (эпифиз, пинеальная железа), разрешенный к медицинскому применению с 1990 года и назначается при климактерических миокардиопатиях [8]. Оба препарата – российские разработки.
Эпиталон способен удлинять теломеры, индуцируя активность теломеразы [17-19]. В экспериментах на теломераз-позитивной клеточной линии HeLa и первичной культуре фибробластов легких плода человека 602/17 начиная с 27-го пересева клетки обрабатывали Эпиталоном (конц. 0,05 мкг/мл) четыре дня, после чего анализировали с помощью иммуногистохимического окрашивания с использованием мышиных моноклональных антител к субъединице теломеразы человека NCL-hTERT [18]. В качестве второго антитела служил стандартный набор пероксидазы ABC. Активность теломеразы оценивали в соответствии с протоколом TRAP, и среднюю длину теломер в отдельных клетках определяли проточной цитофлуорометрией (flow-FISH) и флуоресцентной гибридизацией in situ. Иммуногистохимия показала интенсивное окрашивание теломераз-положительных клеток HeLa, используемых как позитивный контроль, и фибробластов плода, обработанных Эпиталоном. Окрашивание не наблюдалось в интактных (контрольных) фибробластах. ПЦР-анализ не обнаружил активности теломеразы в интактных фибробластах, тогда как клетки HeLa и фибробласты, подвергшихся воздействию Эпиталона, ее проявляли. Флуоресцентная гибридизация и flow-FISH продемонстрировали увеличение средней и максимальной длины теломер в клетках после Эпиталона. Эти результаты показывают, что Эпиталон может усиливать выработку ферментного компонента теломеразы, активность теломеразы (на ~33,3%), а длина теломер увеличивалась в 2,4 раза. Это говорит о преодолении предела Хейфлика (в норме ~50 делений), что объясняет геропротекторное действие Эпиталона и указывает на возможность продления продолжительности жизни популяция клеток и всего организма.
В недавней работе авторы [19] изучили действие Эпиталона на изменение митотического индекса PHA-стимулированных лимфоцитов крови и длины теломер у мужчин двух возрастных групп (5 чел. 18–22 года и 6 чел. 49–54 года). У молодых людей значимых изменений митотического индексе не было обнаружено, у лиц среднего возраста снижение этого показателя было выявлено в единичном случае. Относительную длину теломер оценивали флуоресцентной гибридизацией с ДНК-зондами, специфичными для теломер. Лимфоциты культивировали с использованием стандартного полумикрометода со стимуляцией фитогемагглютинином (PHA). На 48-й час Эпиталон добавляли во флаконы с культурой до конечной концентрации 1000 нг/мл. Препарат не был добавлен в контрольный флакон. На 71-й час во флаконы вводили 1% колхицина. Для оценки относительной длины теломер метафазных хромосом проводилась FISH со специфичными ДНК-зондами. После инкубации с Эпиталоном удлинение длины теломер были обнаружены у семи человек (3 – у молодых и 4 – у лиц среднего возраста). Значительное удлинение после применения Эпиталона было у двух молодых лиц (41%, 55%) и трех – среднего возраста (156%, 18%, 76%). У двух человек из разных групп длина теломер уменьшилась (на 15% и 37%). Была отмечена склонность к нормализации длины теломер: длина увеличилась у индивидуумов с первоначально низкой длиной в сравнении со средним значением в когорте и уменьшился у мимевших первоначально более длинные теломеры [19].

Биологическая активность пептидов эпифиза обусловлена тем, что шишковидное тело секретирует множество регуляторов, нейромедиаторов и гормонов. Одним из них является мелатонин, который регулирует сон человека в суточных и сезонных циклах и его фазы [9]. По сути, выработка мелатонина – это главная и наиболее важная функция эпифиза, так как данный нейромедиатор проявляет еще и выраженные иммуностимулирующие и антиоксидантные эффекты [10]. Его секреция стимулируется в темноте и подавляется под действием освещения [9]. Предшественником мелатонина является 5-гидрокситриптамин (5-HT, серотонин), который также представляет собой нейромедиатор, участвующий в передаче сигналов в ЦНС и вырабатываемый в головном мозге [11]. Он действует на когнитивные функции и эмоциональное состояние, двигательную активность и тонус сосудов [12, 13].
Мелатонин, главный регулятор хронобиологической и эндокринной физиологии человека, широко известен своим антиоксидантными, иммуномодулирующими, антипролиферативными, онкостатическими и эндокринно-модулирующими эффектами. Многие исследователи полагают, что мелатонин является антивозрастным средством, которое может уменьшить последствия старения и продлить продолжительность жизни [16]. Важность мелатонина в старении обсуждается, в основном, в четырех аспектах. Во-первых, мелатонин является ключевой молекулой, участвующей в поддержании амплитуд циркадных ритмов. Любое нарушение циркадных часов связано со старением и заболеваемостью. Таким образом, «сброс» циркадных часов может быть хорошим вариантом терапии для здорового протекания старения и долголетия. Второй аспект относится к метаболическому действию мелатонина, опосредованному модуляцией важных сигнальных путей, таких как Sirtuin 1 (супрессор старения), AMP-зависимой протеинкиназы (AMPK), фосфатидилинозитол 3-киназы (PI3K) и Akt. В-третьих, мелатонин также участвует в регуляции синтеза митохондриальных белков, улучшении дыхательной эффективности, предотвращении перекисного окисления кардиолипина и связанных с ним антиапоптотических эффектах. Открытие синтеза мелатонина в митохондриях может быть критическим аспектом в контексте старения. Защитные эффекты мелатонина приводят к значительному угнетению митохондриального апоптоза и митофагии. Мелатонин также оказывает мощное иммуномодулирующее действие. Эта функция имеет отношение к ремоделированию иммунитета во время старения и является важным в целом. Кроме того, мелатонин может активировать антиоксидантные ферменты, улучшать образование и доступность восстановленного глутатиона и модулировать метаболизм оксида азота (NO) [16]. Мелатонин в крови обычно определяют методом иммуноферментного анализа (ИФА). Пониженная концентрация нейромедиатора ассоциируется с нарушениями сна, циркадных ритмов и ухудшением состояния иммунной системы. В продаже встречаются различные наборы реагентов по относительно невысоким ценам. Так, «ELISA Kit for Melatonin (MT)» на 96 тестов от Cloud-Clone Corp. стоит примерно 70 тыс. руб., а разовый анализ в российских клиниках стоит от 2,5 тыс. руб.
Эпиталон (даже будучи в составе эпиталамина) примечателен тем, что способен стимулировать секрецию мелатонина, что было показано в экспериментах in vivo на приматах и человеке [14, 15]. Уровень мелатонина является важным показателем и связан со старением организма [16].
В работе [14] были изучены антиоксидантные эффекты мелатонина и эпиталамина и подавление свободнорадикального окисления in vivo на грызунах (по 0,5 мг подкожно, курс – 5 дней). Другая группа получала мелатонин (вода перорально, 20 мг/л). Эпиталон – аналогично, в дозировке по 0,1 мкг. Эпиталамин существенно уменьшал (в 3,4 раза) хемолюминесценцию сыворотки крови и четырехкратно понижал перекисное окисление липидов. При этом на 35% возросла общая антиоксидантная активность. После недельного курса эпиталамина активность супероксиддисмутазы (СОД) возросла на ~20%; мелатонин подавил активность на ~37%. СОД в крови крыс после курса эпиталамина на 75% превысил таковой для группы, которой давали раствор мелатонина [14]. Таким образом, антиоксидантные свойства Эпиталона и эпиталамина превосходят действие мелатонина.
В норме, концентрация мелатонина у приматов и человека демонстрирует выраженную зависимость от циркадного ритма, достигая максимума ночью (до 130 пг/мл у здоровых молодых людей и до 40 пг/мл – у пожилых) с дневным минимумом (30 пг/мл и 10 пг/мл у молодых и пожилых, соответственно) [15]. Такая зависимость от возраста обусловлена снижением функциональной активности эпифиза у ~71% людей. Авторы работы [15] исследовали секрецию мелатонина на молодых и старых самках макак (Macaca mulatta). Животных разделили на группы по 4 молодых и старых особей и на протяжении 10 дней вводили эпиталамин в дозе 5 мг/сутки/животное внутримышечно. Контроль – плацебо (инъекции солевого раствора). Двум группам по 6 молодых и старых особей тот же период вводили Эпиталон по 10 мкг/сутки/животное внутримышечно. В плазме крови определяли концентрацию мелатонина иммуноферментным методом (реактивы Melatonin-ELISA», ФРГ) с предварительною очисткой пробы на хроматографических колонках, и таким образом оценивали среднесуточную концентрацию мелатонина (пг/мл) и амплитуду суточного ритма. Также, клинические исследования проводили на молодых (15 человек, 20–34 лет) и пожилых (49 человек, 60–79 лет) людях [15]. В плазме уровень мелатонина измеряли с помощью стандартных радиоиммунных наборов компании DPС (США), причем образцы венозной крови забирали через предварительно установленный миникатетер в течение суток. Добровольцев рандомизировали в три однотипные группы: группа 1 (15 чел.) – эпиталамин по 10 мг внутримышечно раз в трое суток, 5 введений инъекций, суммарная доза 50 мг, группа 2 (15 чел.) — Эпиталон по 0,01 мг ежедневно, 10 инъекций внутримышечно, курсовая доза 0,1 мг, группа 3 (10 чел.) — плацебо (физраствор, 10 сут ежедневно). В результате было обнаружено, что Эпиталон и эпиталамин на молодых животных практически не оказывал влияния, тогда как у старых особей, получавших Эпиталон, через 10 дней наблюдалось возрастание концентрации вечернего мелатонина до 1,8 раз, по сравнению с исходным уровнем и с контролем, и достигали уровня нейромедиатора и его суточного ритма, характерного для молодых обезьян. Эпиталамин вызвал увеличение концентрации мелатонина вечером и ночью, а амплитуда циркадианного ритма возросла двукратно. Эпиталон, при сопоставлении эффекта и дозировки, продемонстрировал пятисоткратное превосходство биологической активности над эпиталамином. Испытания на добровольцах показали, что у пожилых со сниженной выработкой мелатонина эпиталамин увеличил ночную концентрацию мелатонина более чем двукратно. Применение Эпиталона оказалось аналогичным по действию, но повышалась и вечерняя концентрация нейромедиатора [15].
Очевидно, что контроль длины теломер и уровня мелатонина в крови могут быть важными прогностическими показателями. Эти данные можно использовать для прогнозирования продолжительности жизни и корректировки лечащим врачом текущей терапии. Препараты, позитивно регулирующие концентрацию мелатонина и длину теломер, возможно, найдут применение в геронтологии и в терапевтических подходах, направленных на продление жизни и улучшение ее качества в пожилом и преклонном возрасте.
1. Marotta, F., Aging Interventions and Therapies, Editor: Suresh I.S. Rattan; World Scientific, Singapore, 2005; pp 418; hard back; price USD85; ISBN 981256084X. Biogerontology, 2005. 6(5): p. 369-370. DOI: 10.1007/s10522-005-4812-4.
2. Cawthon, R.M., Telomere measurement by quantitative PCR. Nucleic acids research, 2002. 30(10): p. e47-e47. DOI: 10.1093/nar/30.10.e47.
3. Baird, D.M., Rowson, J., Wynford-Thomas, D., Kipling, D., Extensive allelic variation and ultrashort telomeres in senescent human cells. Nature Genetics, 2003. 33(2): p. 203-207. DOI: 10.1038/ng1084.
4. Montpetit, A.J., Alhareeri, A.A., Montpetit, M., Starkweather, A.R., Elmore, L.W., Filler, K., Mohanraj, L., Burton, C.W., Menzies, V.S., Lyon, D.E., Jackson-Cook, C.K., Telomere length: a review of methods for measurement. Nurs Res, 2014. 63(4): p. 289-299. DOI: 10.1097/NNR.0000000000000037.
5. Khavinson, V.K., Peptides and ageing. Neuroendocrinology Letters, 2002. 23(Suppl 3): p. 11-144.
6. Khavinson, V.K., Izmaylov, D.M., Obukhova, L.K., Malinin, V.V., Effect of epitalon on the lifespan increase in Drosophila melanogaster. Mechanisms of Ageing and Development, 2000. 120(1): p. 141-149. DOI: https://doi.org/10.1016/S0047-6374(00)00217-7.
7. Anisimov, V.N., Sredstva profilaktiki prezhdevremennogo stareniya (geroprotektory) [Prevention of premature aging (geroprotectors)]. Uspekhi Gerontologii [Advances in Gerontology], 2000. 4: p. 275-277.
8. Приказ «О разрешении к медицинскому применению» от 19 июня 1990 года N 250. Министерство Здравоохранения СССР
9. Lowrey, P.L., Takahashi, J.S., Genetics of the mammalian circadian system: Photic Entrainment, Circadian Pacemaker Mechanisms, and Posttranslational Regulation. Annual Review of Genetics, 2000. 34(1): p. 533-562. DOI: 10.1146/annurev.genet.34.1.533.
10. Espino, J., Pariente, J.A., Rodriguez, A.B., Oxidative Stress and Immunosenescence: Therapeutic Effects of Melatonin. Oxidative Medicine and Cellular Longevity, 2012. 2012: p. 9. DOI: 10.1155/2012/670294.
11. Frazer, A., G., H.J., Understanding the neuroanatomical organization of serotonergic cells in the brain provides insight into the functions of this neurotransmitter, in Basic neurochemistry: molecular, cellular, medical aspects, G.J. Siegel, Agranoff, B.W., Fisher, S.K., Albers, R.W., Uhler, M.D., Editors. 1999, Lippincott Williams & Wilkins. p. 264-268.
12. Chilmonczyk, Z., Bojarski, A., Pilc, A., Sylte, I., Functional selectivity and antidepressant activity of serotonin 1A receptor ligands. International journal of molecular sciences, 2015. 16(8): p. 18474-18506.
13. Blier, P., El Mansari, M., Serotonin and beyond: therapeutics for major depression. Philosophical Transactions of the Royal Society B: Biological Sciences, 2013. 368(1615): p. 20120536. DOI: 10.1098/rstb.2012.0536.
14. Kozina, L.S., Arutjunyan, A.V., Khavinson, V.K., Antioxidant properties of geroprotective peptides of the pineal gland. Archives of Gerontology and Geriatrics, 2007. 44: p. 213-216. DOI: 10.1016/j.archger.2007.01.029.
15. Коркушко, О.В., Лапин, Б.А., Гончарова, Н.Д., Хавинсон, В.Х., Шатило, В.Б., Венгерин, А.А., Антонюк-Щеглова, И.А., Магдич, Л.В., Нормализующее влияние пептидов эпифиза на суточный ритм мелатонина у старых обезьян и людей пожилого возраста. Успехи геронтологии, 2007. 20(1): p. 74-85.
16. Majidinia, M., Reiter, R.J., Shakouri, S.K., Yousefi, B., The role of melatonin, a multitasking molecule, in retarding the processes of ageing. Ageing Research Reviews, 2018. 47: p. 198-213. DOI: https://doi.org/10.1016/j.arr.2018.07.010.
17. Anisimov, V.N., Khavinson, V.K., Mikhailova, O.N., Biogerontology in Russia: from past to future. Biogerontology, 2011. 12(1): p. 47-60. DOI: 10.1007/s10522-010-9307-2.
18. Khavinson, V.K., Bondarev, I.E., Butyugov, A.A., Epithalon Peptide Induces Telomerase Activity and Telomere Elongation in Human Somatic Cells. Bulletin of Experimental Biology and Medicine, 2003. 135(6): p. 590-592. DOI: 10.1023/A:1025493705728.
19. Khavinson, V.K., Pendina, A.A., Efimova, O.A., Tikhonov, A.V., Koltsova, A.S., Krapivin, M.I., Petrovskaia-Kaminskaia, A.V., Petrova, L.I., Lin’kova, N.S., Baranov, V.S., Effect of Peptide AEDG on Telomere Length and Mitotic Index of PHA-Stimulated Human Blood Lymphocytes. Bulletin of Experimental Biology and Medicine, 2019. 168(1): p. 141-144. DOI: 10.1007/s10517-019-04664-0.




{kind=link}
{kind=link}
{kind=link}
{kind=link}