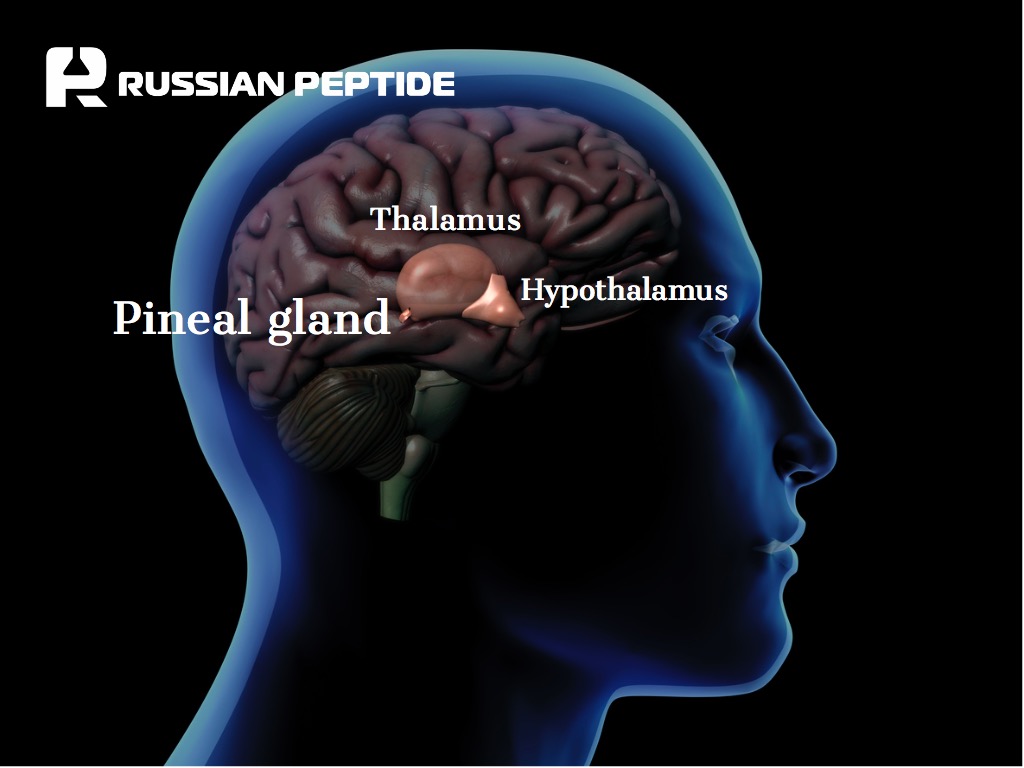
Пептиды шишковидной железы
04.06.2020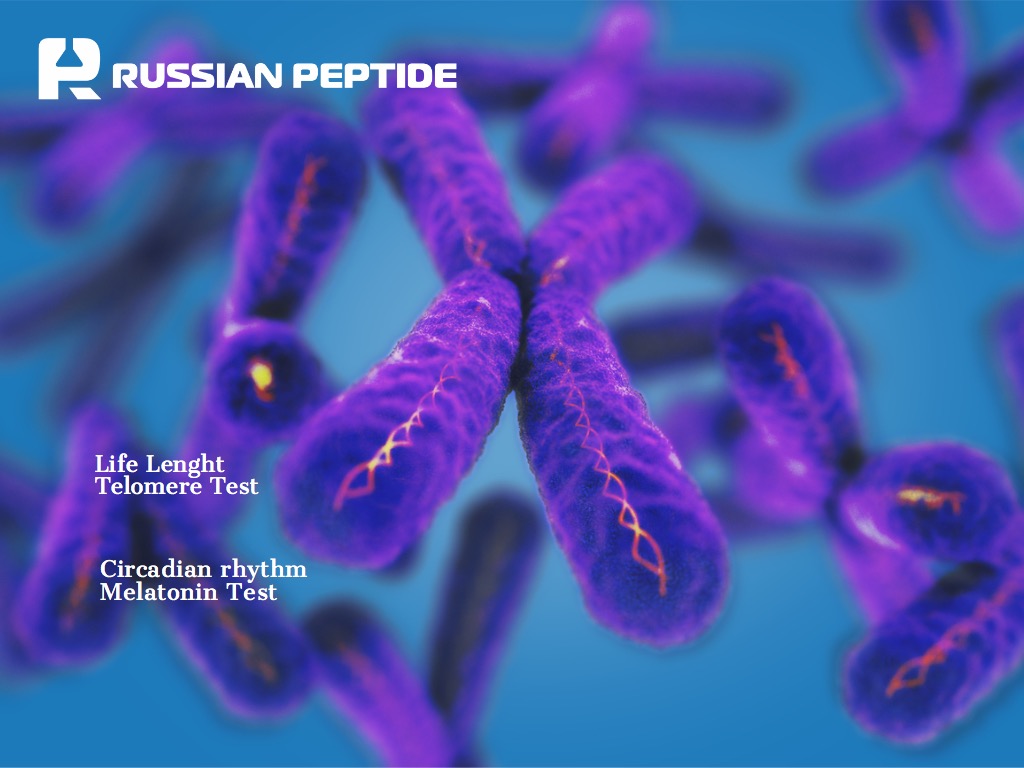
Эпиталон. Тест на теломеры и мелатонин
14.06.2020
HEP1, известный также как Гепон (Gepon, Hepon, Human Ezrin Peptide One, Human Ezrin Peptide 324-337) и зарегистрированный на территории России в качестве иммуномодулирующего лекарственного средства с противовирусным эффектом (ЛСР-001794/07) представляет собой синтетический пептид, имитирующий 14 аминокислот из последовательности белка TEKKRRETVEREKE в α-домене человеческого эзрина, многофункционального мембраносвязанного белка [1]. За прошедшие 25 лет HEP1 показал свою безопасность для применения человеком, что было подтверждено исследованиями, проведенными в Германии, Великобритании и России. HEP1 обладает множественной активностью, так как он является модулятором клеточной сигнальной системы. Это селективный противовирусный промотор Т-клеточных и В-клеточных иммунных ответов, HEP1 стимулирует фибробласты, способствуя восстановлению тканей, в то же время подавляя воспалительные реакции за счет повышения уровня интерлейкина IL-10, что приводит к снижению выработки воспалительных цитокинов IL-1-β, IL-6, IL-8 и TNF-α. Вирус SARS-CoV-2 приводит к инфекционному заболеванию COVID-19, которое примерно в 1% случаев связано с тяжелыми острыми и хроническими воспалительными процессами. В пилотных клинических исследованиях было обнаружено, что HEP1 обладает быстрым действием и уменьшает воспаление, лихорадку и кашель у пациентов с COVID-19 в течение нескольких часов, а также полностью устраняет SARS-CoV-2 в течение нескольких дней.
Исторически, разработка пептидных препаратов на основе эзрина началась с поиска средства от ВИЧ-инфекции. В 1993 году в ходе совместной исследовательской программы по эпидемии ВИЧ-инфекции в Лондоне, посвященной поиску потенциальной вакцины на основе пептидов против ВИЧ, д-р Руперт Холмс (Dr Rupert Holms) обнаружил неожиданную гомологию между карбокси-С-концом белка gp120 ВИЧ и центральным α-доменом человеческого эзрина (Табл. 1). Отметим, что д-р Р. Холмс является соучредителем и председателем московской фармацевтической компании Nearmedic Group, со штатом около тысячи человек и объемом продаж порядка 100 млн. евро. Он является основным изобретателем фармацевтических пептидных препаратов на основе эзрина [2].
С-конец белка gp120 ВИЧ и центральный α-домен эзрина являются α-спиралевидными структурами и они высокозаряжены. Совпадение зарядов аминокислот боковой цепи и вторичной структуры позволяет предположить, что последовательности gp120 и эзрина человека имеют сходную функцию, причем уровень мутаций на С-конце gp120 близок к нулю. В 1987 году было обнаружено, что мышиное моноклональное антитело под названием M38, которое было получено против супернатанта ВИЧ-1 из инфицированных Т-клеток, имело серологическую перекрестную реактивность между гликопротеином gp120 ВИЧ-1 и белком на поверхности клеток человека, который экспрессируется на небольшой фракции моноцитов в периферической крови и в лимфатических узлах, и имеет молекулярную массу 80 кДа. Позднее пептидное картирование показало, что М38 связывается и с синтетическим gp120-мимиком (см. Табл. 1). Связанный с мембраной эзрин экспрессируется в активированных моноцитах во время острых воспалительных реакций и необходим для миграции в сторону инфицированной или поврежденной ткани.

В 1997 году стало известно, что Mab 670-D, моноклональное антитело, активное по отношению к последовательности TKAKRR на С-конце gp120, эффективно блокировало опосредованное gp120 слияние и проникновение ВИЧ-1 в клетки. Дальнейшая работа над концевым пептидным фрагментом gp120 показала, что, из-за его высокой аффинности к рецепторным/транспортным белкам клеточной поверхности, он блокирует gp120-опосредованное слияние клетка-клетка и вирус-клеточное слияние.
Эзрин (также известен под названием виллин-2 или цитовиллин) является членом эволюционно консервативного семейства белков Ezrin/Radixin/Moesin (ERM). В его состав входит 586 аминокислот, масса составляет 69,4 кДа. Эзрин имеет различные функциональные формы и степени фосфорилирования, которые связаны с упорядочиванием рецепторов, сигнальных белков и актинового цитоскелета в активные комплексы на клеточной мембране. Эзрин, весьма вероятно, является рецептором слияния для короновируса SARS-CoV-2.

Рисунок 1 – Формы эзрина
Эзрин имеет три основные и несколько переходных форм (Рис. 1). Существует компактная «неактивная» глобулярная конформация, которая растворима в цитоплазме. Известна полностью развернутая «активная» конформация, связанная с богатыми PIP2 областями внутренней поверхности клеточной мембраны. Накапливаются данные о промежуточной трансмембранной «рецепторной» форме эзрина, в которой С-концевой и FERM домены остаются на внутренней поверхности клеточной мембраны, тогда как α-домен образует рецепторную структуру, несущую электростатический заряд и выходящую на наружную поверхность клеточной мембраны (Рис. 1).
Белковые комплексы на основе эзрина различаются по функции в зависимости от компонентов и состояния фосфорилирования эзрина. Эзрин контролирует организацию и интернализацию рецепторов, передачу сигналов, а также модулирует молекулы клеточной адгезии и связывание интегрина, кадгерина и ICAM.
Эзрин опосредует отрицательную регуляцию сигнального пути рецепторов Т-клеток, каскада ras>raf>MEK>ERK1/2 и p38MAPK, как и передачи сигналов интерлейкина-2. Эзрин также положительно регулирует передачу сигналов, опосредованную фосфатидилинозитом, посредством передачи сигналов PI3K> Akt> mTOR и протеинкиназы А. Эзрин контролирует связывание АТФазы, клеточный ответ на связывание цАМФ и белка S100-Ca2+. Эзрин обеспечивает положительную регуляцию экспрессии генов, но отрицательную регуляцию транскрипции РНК-полимеразой II. Эзрин контролирует клеточную структуру и подвижность клеток посредством регуляции формы клеток путем позитивной регуляции расположения белков на плазматической мембране или в кортикальном слое клетки. Эзрин также управляет прикреплением цитоскелета к плазматической мембране, установлением и поддержанием полярности апикальных/базальных клеток, филоподиевой сборкой, локализацией центросом. Эзрин также регулирует сборку органелл, реорганизацию актинового цитоскелета, закрепление мембраны, организацию астральных и корковых микротрубочек и управление аксонами. Эзрин также обеспечивает позитивную регуляцию транспортировки ранний и поздних эндосом, а также катаболических процессов в клеточном белке. В кишечнике эзрин контролирует целостность слизистой оболочки и ее восстановление, сборку микроворсинок и регулирование их длины, абсорбцию D-глюкозы и позитивную регуляцию секреции белка. Эзрин контролирует мембранный транспорт через регуляторы Na+/H+-обмена NHERF1 и PKA, поддерживает фосфатный гомеостаз и передачу сигналов цАМФ. Эзрин также регулирует миграцию макрофагов, фибробластов и дендритных клеток, управляет эндотелиальным барьером и положительной регуляцией роста многоклеточного организма.
Различные рецепторные, сигнальные и структурные белки имеют специфические сайты связывания с эзрином (Рис. 2). Эзрин имеет три домена: трехлепестковый глобулярный домен FERM (F1–F3 и концевой транс-FERM), гибкий α-домен (αhelixA1, αhelixA2, αhelixB1, αhelixB2, αhelixC1, αhelixC2, α-trans-pro и α-flap), которые могут принимать различные формы путем свертывания или разворачивания структуры α-спирали и небольшого глобулярного C-терминального домена (C-ERMAD). С FERM-доменом эзрина связываются следующие белки: цитоплазматические концевые сегменты рецепторов клеточной поверхности, интегрин, кадгерин, ICAM-1, ICAM-2, VCAM-1, EGF-R, CD43, CD44, CD95; сигнальные белки, ras, ras-подобная GTPase Rho, ингибитор диссоциации Rho-GDP [Rho-GDI] и Dbl, Grb2, SOS, FAK, PTP. Линкерный и насосный белки EPB50 и NHERF связываются с доменом FERM, а также с мембранным фосфолипидом PIP2. Сигнальные киназы PKC, p85-PI3K и KA связываются с α-доменом эзрина. 
Цитоскелетный белок актин связывается с С-концевым доменом эзрина.

Рисунок 2 – Связывание белков с эзрином
Коронавирус, ВИЧ И ВГС используют эзрин для проникновения в клетку
Оказалось, что рецептор/транспортный белок клеточной поверхности представляет собой новую конформацию человеческого эзрина, которая выходит на поверхность клеток и имеет схожую с застежкой-молнией лакуну, состоящую из комплементарных заряженных боковых цепей на двух смежных α-спиралях (Рис. 3). После связывания с CD4, C-конец gp120 взаимодействует с данной лакуной, «расстегивает» молнию, вызывая конформационное изменение в белке, которое запускает слияние вируса с клеткой и различные последующие события. HEP1 взаимодействует с той же расщепленной структурой в экспрессируемом эзрине на клеточной поверхности, что запускает интернализацию эзрина и затем – специфические биологические процессы в клетках. В настоящее время известно, что не только ВИЧ использует эзрин для проникновения в клетки, но и вирус гепатита С (ВГС), и коронавирус SARS. Для ВИЧ и SARS-CoV-2 новый белковый рецептор/транспортер, вероятно, является трансмембранной «рецепторной» конформацией человеческого эзрина.

Рисунок 3 – Эзрин, интернализация В-клеточных рецепторов и слияние вирусов
Конформационное переключение эзрина регулирует динамику В-клеточного рецептора (BCR) и может быть ключевым для слияния коронавируса с клеткой. После стимуляции BCR с помощью анти-IgM, эзрин фосфорилируется по Y353, который находится рядом с сайтом связывания HEP1. Y353-фосфорилированный эзрин затем совместно локализуется с BCR в течение нескольких минут с последующим эндоцитозом BCR из мембраны в клетку. После, эзрин совместно участвует с эндоцитозированными BCR через ранние и поздние эндосомы. Эзрин-зависимый эндоцитоз, наблюдаемый в В-клетках, может быть распространенным механизмом, используемым ВИЧ, ВГС и коронавирусами для слияния с мембраной клетки-мишени и проникновения во многие типы клеток.
HEP1 быстро уменьшает воспалительные процессы при COVID-19. Инфекция SARS-CoV-19 усиливает секрецию провоспалительных цитокинов IL-1, IL-6, IL-8, TNF-α и вызывает тяжелые симптомы COVID-19. Избыточная экспрессия IL-6, вероятно, является основной причиной повреждения легких при COVID-19.
Провоспалительные цитокины
Интерлейкины-1 (IL-1α и IL-1β) синтезируются множеством типов клеток по всему организму, включая моноциты, макрофаги, нейтрофилы, B-клетки, дендритные клетки, NK-клетки, T-клетки, гепатоциты и макрофаги тканей. IL-1β оказывает стимулирующее действие на CD4+ Т-клетки и способствует дифференцировке Т-хелперов.
Интерлейкин-6 (IL-6) в ранних исследованиях был идентифицирован как стимулятор секреции IgG линиями В-лимфобластоидных клеток и как фактор роста В-клеток. IL-6 выделяется Т-клетками.
Интерлейкин-8 (IL-8 или CXCL8) является одним из наиболее изученных провоспалительных хемокинов, поскольку он встречается в высоких концентрациях в плазме во время воспаления (1–100 нг/мл). IL-8 продуцируется макрофагами, а его основной мишенью являются нейтрофилы.
Фактор некроза опухоли (TNF-α) производится моноцитами и макрофагами, действует на другие макрофаги, вызывая фагоцитарную активацию и эндотоксический шок. TNF-α связывается с доменами CRD тримеров рецептора TNF (TNFR) на внеклеточной поверхности клеток-мишеней.
Интерлейкин 10 (IL-10) является главным регулятором иммунной системы. IL-10 может продуцироваться практически всеми иммунными клетками, и, в свою очередь, IL-10 способен модулировать ответ этих клеток. Основная функция IL-10 заключается в предотвращении иммунопатологии путем подавления чрезмерной экспрессии провоспалительных цитокинов. Однако система регулирования IL -10 сложна и, отчасти, парадоксальна. IL-10 настолько важен для регуляции иммунного ответа, что вирусы эволюционировали так, чтобы использовать регуляцию IL-10 для обхода иммунитета и появления хронических/латентных инфекций. SARS-CoV-2 ингибирует регуляторную систему IL-10. Регуляция секреции IL-10 и влияние IL-10 на клетки-мишени обеспечивает объяснение быстрого системного противовоспалительного эффекта, наблюдаемого при лечении клинического воспаления с помощью HEP1 (Гепон) при COVID-19 и других заболеваниях. IL-10, как правило, подавляет воспалительные реакции в конечной фазе иммунологического контроля вирусной инфекции. Однако SARS-CoV-19 мешает контролю IL-10, ингибируя его экспрессию.
Секреция IL-10 регулируется множеством сигнальных путей, включая p38, PI3K, ERK и NF-κB, которые инициируются в мембранно-ассоциированных эзрин-белковых комплексах. Основным путем является TLR> MyD88. Кроме того, цАМФ-протеинкиназа A (PKA) связывается с α-доменом активированного удлиненного эзрина с образованием белкового комплекса; передача сигналов вниз увеличивает экспрессию IL-10. Связанный с эзрином путь ras>raf>MEK>ERK (MAPK) также способствует как инициации транскрипции IL-10, так и посттранскрипционной регуляции, чтобы стимулировать и поддерживать выработку IL-10. Механизмы, регулирующие выработку IL-10 в лейкоцитах (WBC), полностью не изучены. Однако эзрин-зависимый каскад TLR>TRIF>TANK-TBK1>IRF3> NF-kB особенно важен в экстренном контроле воспаления. Сверх- экспрессия IL-10 контролируется эзрином, который, будучи полностью активным, ограничивает избыточную выработку IL-10 путем ингибирования пути TLR>TRIF>TANK-TBK1>IRF3>NF-kB. Нокдаун-мыши, у которых отсутствует функция MyD88 или TRIF, быстро гибнут после заражения SARS-CoV.
Высокие уровни IL-6 в сыворотке связаны с одышкой (затрудненное дыхание), рвотой и низким насыщением кислородом, что часто встречается при тяжелом течении COVID-19. IL-6 является наиболее важным цитокином в определении тяжести заболевания у пациентов с синдромом системного воспалительного ответа.
На ранних стадиях пневмонии, во время реакции острой фазы, альвеолярные макрофаги в легких продуцируют различные воспалительные цитокины и хемокины: IL-1, IL-6, IL-8, TNF-α и интерферон-гамма (IFN-γ). Избыток IL-6 связан с тяжелой пневмонией и повреждением легких при COVID-19. Отношение сывороточного IL-6 к IL-10 выше пятикратного является признаком тяжелой пневмонии. Напротив, модуляция воспалительного ответа в дыхательных путях и восстановление после пневмонии связано с экспрессией противовоспалительных цитокинов IL-10 и IL-12.
Противовоспалительная активность HEP1, вероятно, обусловлена его действием на конформацию рецептора эзрина, которая индуцирует изменения в конформационных состояниях и степени фосфорилирования в трех основных клеточных пулах, что приводит к повышению выработки IL-10. Одна из гипотез состоит в том, что HEP1 увеличивает обновление эзрина, приводя к более низким относительным количествам активной конформации эзрина и к подавлению сигнального пути TLR4-TRIF-IRF3, вызывая увеличение концентрации IL-10 в сыворотке. Это изменение, в свою очередь, воздействует на клетки, ингибируя экспрессию IL-1, IL-6, IL-8 и TNF-α, и быстро уменьшая клиническое воспаление.

Рисунок 4 – Действие HEP1 на иммунный ответ и цитокины (пептиды см. Табл. 1)
Пептиды на основе эзрина активируют эзрин человека, интенсифицируя различные реакции адаптивной иммунной защиты, запрограммированные Т-клетками и В-клетками. Эзрин также регулирует выработку IL-10, главного регулятора воспалительных цитокинов. Основная функция IL-10 заключается в предотвращении иммунопатологии при воспалительных реакциях, которая является серьезной проблемой при COVID-19. HEP1 модулирует фосфорилированный эзрин для усиления передачи сигналов IL-10, одновременно уравновешивая воспаление и иммунный ответ во время прогрессирования заболевания COVID-19. HEP1 также активирует фосфорилированный эзрин на внутренней поверхности клеточной мембраны, предотвращая слияние вируса и проникновение путем эндоцитоза. Кроме того, HEP1 активирует соответствующие антивирусные ответы T- и B- клеток.
Таким образом, в основе действия пептидной терапии COVID-19 лежат противовоспалительная активность, антивирусное действие (подавление слияния) и усиление адаптивного противовирусного иммунного ответа. Кроме того, пептиды на основе эзрина безопасны, и побочные эффекты для них не описаны.
С другими свойствами и описаниями клинических исследований HEP1 можно ознакомиться в предыдущем обзоре. Синтетические аналоги Гепона рассмотрены нами в недавнем обзоре.
Текст настоящего обзора основан на находящихся в печати материалах «Review of Mechanism of Action of Ezrin Peptides. Relevance to COVID-19 Treatment», Dr. Rupert Holms. Графики и иллюстрации адаптированы из указанной работы.
1. GB 2290293 A, 1994.
2. US 9682140 B2, 2017.
3. GB 0301879 D0, 2003.
4. US 5773573 A, 1998.
5. US 6849596 B1, 2005.




{kind=link}
{kind=link}
{kind=link}
{kind=link}