
Роль кисспептина в регуляции репродуктивных функций и метаболических процессов
14.10.2022
β-Лактолин: память и внимание, депрессия и утомляемость
24.10.2022
L-Метионин – незаменимая протеиногенная аминокислота. Нехватка незаменимых аминокислот может провоцировать нарушения метаболизма, однако в некоторых случаях пониженное потребление метионина является полезным. В обзоре рассмотрено положительное влияние ограничения метионина в диете на метилирование ДНК, окислительные процессы, ожирение, продолжительность жизни, состояние пациентов с онкологическими заболеваниями, а также негативные эффекты на рост организма и плотность костей.
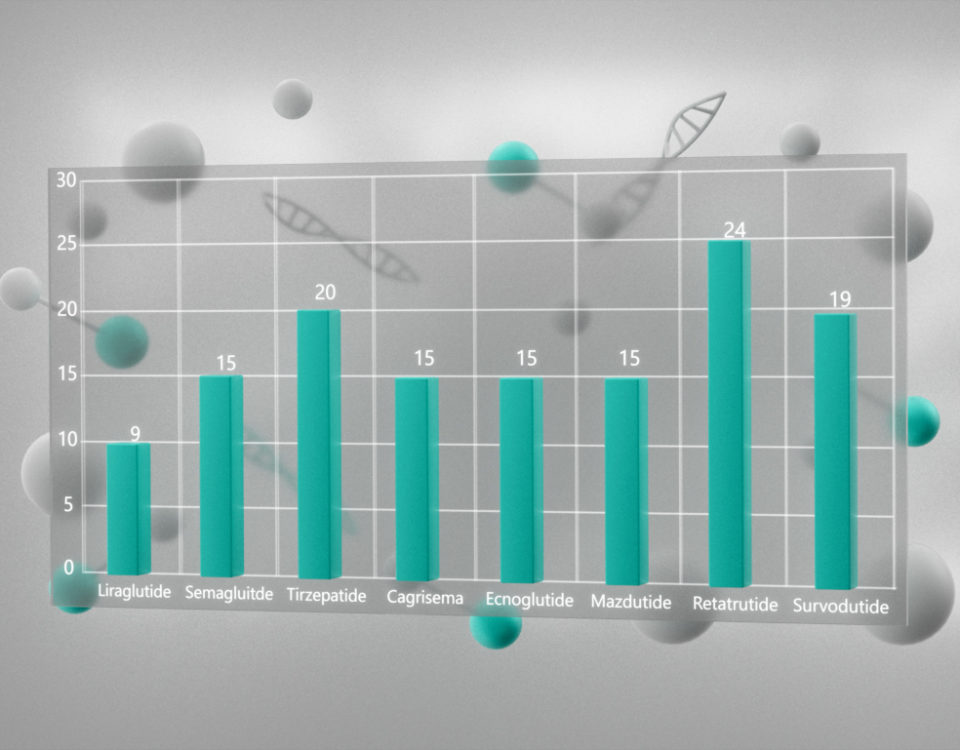
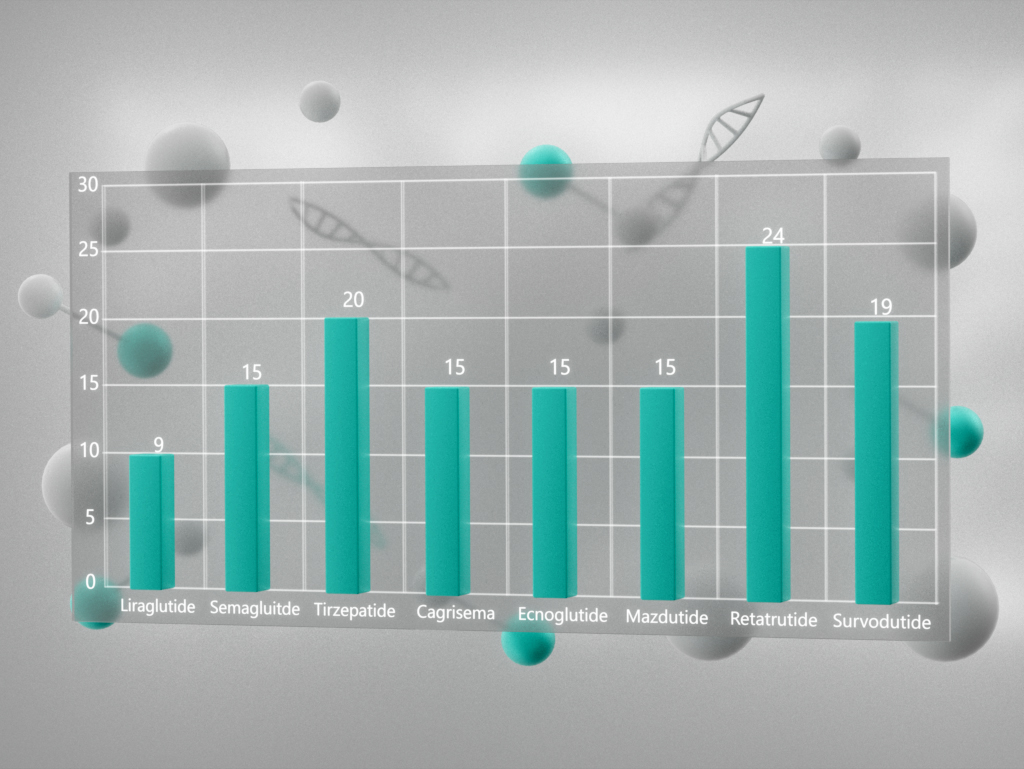
L-Метионин (L-Methionine, Met, M, (S)-2-амино-4-(метилтио)масляная кислота, (S)-2-амино-4-(метилтио)бутановая кислота, C5H11NO2S, CAS 63-68-3) – протеиногенная незаменимая аминокислота, выделенная впервые в 1921 году. Поскольку незаменимые аминокислоты не синтезируются в организме человека, источником метионина служат белки, поступающие с пищей. Ниже приведено среднее содержание метионина в различных продуктах (Рис. 1).

Рисунок 1 – Содержание метионина в различных пищевых источниках (столбцы представляют собой среднюю концентрацию; цифры внутри столбцов – количество источников, на основании которых выведено среднее значение) [1]
Животные и взрослые люди превращают D-метионин в L-метионин путем трансаминирования через α-кетокислоту, поэтому D- и L-метионин имеют одинаковую питательную ценность. Синтетический рацемат можно напрямую использовать в качестве пищевой добавки. Поскольку метионин, как и лизин и треонин, коммерчески доступны и недороги, их часто используют в составе кормов. Наиболее экономичным способом производства D,L-метионина является химический процесс на основе акролеина, метилмеркаптана, цианистого водорода и карбоната аммония [2]. β-Метилтиопропионовый альдегид, образованный добавлением метилмеркаптана к акролеину, является промежуточным продуктом, который реагирует с цианистым водородом с образованием α-гидрокси-γ-метилтиобутиронитрила. Обработка карбонатом аммония приводит к 5-(β-метилтиоэтил)гидантоину, который омыляется карбонатом калия с образованием D,L-метионина с выходом до 95% в пересчете на акролеин.
Предпочтительным методом получения L-метионина по-прежнему является ферментативное разделение рацемического N-ацетилметионина с использованием ацилазы из плесневого гриба AspergiLLus oryzae. Производство осуществляется в непрерывно работающем реакторе с неподвижным слоем или ферментативном мембранном реакторе. Альтернативно, L-метионин может быть получен микробиотическим превращением соответствующего 5-замещенного гидантоина: клетки Pseudomonas sp., штамм NS671, преобразуют D-1,5-(2-метилтиоэтил)гидантоин в L-метионин до конечной концентрации 34 г/л и с молярным выходом 93% [2].
Добавление небольших количеств аминокислот для улучшения питательной ценности белков хорошо известно. Как добавки, так и комбинация белков с дополнительными аминокислотами используются для увеличения биологической ценности. Обычно в пище запас хотя бы одной из незаменимых аминокислот ниже потребности. Эта ограничивающая аминокислота определяет, какой процент белка (или, точнее, его аминокислот) можно использовать для удовлетворения потребностей организма. В большинстве случаев первой ограничивающей аминокислотой является метионин. Как правило, животный белок содержит незаменимые аминокислоты в большем количестве и в более благоприятном соотношении, чем растительный. Так, метионин является первой ограничивающей аминокислотой в мясе, молоке, соевых и других бобах [2].
У некоторых младенцев наблюдается несовместимость с лактозой или белками коровьего молока. Пищевые добавки, рекомендованные для этого состояния, часто основаны на очищенном соевом белке, дополненном L-метионином для повышения биологической ценности. Экструдированный соевый белок, который в больших количествах используется в качестве разбавителя мяса и вегетарианского мясозаменителя, обогащают N-ацетил-L-метионином.
Парентеральные инфузии растворами L-аминокислот является хорошо зарекомендовавшим себя компонентом лечебного питания. Стандартный инфузионный раствор содержит восемь классических незаменимых аминокислот, включая метионин, частично заменимые и несколько заменимых аминокислот. L-метионин применяется не только как пищевая добавка, но как липотропное и желчегонное средство. Для пациентов с заболеваниями печени доступны растворы, богатые аминокислотами с разветвленной цепью и бедные ароматическими аминокислотами и метионином. Метионин является донором метильной группы, благодаря чему участвует в эпигенетической регуляции активности генов.
Аминокислоты, белки и их гидролизаты широко используются в средствах для кожи и волос, например, в смягчающих кремах, очищающих лосьонах и шампунях. Метионин всасывается в кожу головы и абсорбируется волокнами волос в виде цистеина, который отвечает за сшивание кератина. Аминокислоты, произведенные путем ферментации, а не извлеченные из животного белка, часто рекламируют как более «натуральные». Серосодержащие аминокислоты оказывают особое нормализующее действие на метаболизм кожи, например, в случаях избыточного производства кожных липидов (себорея), перхоти или угрей. Вещества, используемые для этой цели, включают производные цистеина (например, S—карбоксиметилцистеин), гомоцистеин (2-амино-4-меркаптомасляная кислота) и метионин [2].
Ограничение метионина и продолжительность жизни
Ограничение метионина, как показано на животных моделях, влияет на продолжительность жизни. Снижение с двухнедельного возраста и на протяжении всей жизни концентрации одного пищевого компонента, незаменимой аминокислоты L-метионина, с 0,86% до 0,17% от рациона приводит к увеличению продолжительности жизни самцов крыс на 30% [3]. В эксперименте, когда содержание метионина в рационе было уменьшено, массу глутаминовой кислоты увеличивали пропорционально. При этом, ограничение метионина (MetR – Methionine Restriction) полностью прекратило рост, хотя потребление пищи было больше в зависимости от массы тела. Потребления энергии в раннем возрасте было почти нормальным для животных их размера. Увеличенное потребление энергии крысами, получавшими 0,17% метионина, не привело к повышению скорости их роста, тогда как у крыс, получавших 0,85% метионина, употреблением корма с 0,17% метионина не привело к торможению роста, что указывает на то, что ограничение в еде не было решающим фактором.
В экспериментах на животных было показано, что ограничение потребления метионина, как и диетическое ограничение (DR), увеличивает максимальную продолжительность жизни грызунов [4]. Состав диеты с 40% MetR был близок к составу контрольной пищи, за исключением того, что L-метионин присутствовал в количестве 0,516%, что на 40% ниже, чем в контроле (0,86%). Абсолютное снижение уровня L-метионина на 0,34% в диете с 40% MetR было компенсировано увеличением всех других диетических компонентов пропорционально их присутствию в рационе. Состав контрольной диеты (г/100 г): L-аргинин 1,12; L-лизин 1,44; L-гистидин 0,33; L-лейцин 1,11; L-изолейцин 0,82; L-валин 0,82; L-треонин 0,82; L-триптофан 0,18; L-метионин 0,86; L-глутаминовая кислота 2,7; L-фенилаланин 1,16; L-глицин 2,33; декстрин 5; кукурузный крахмал 31,8; сахароза 31,79; целлюлоза 5; битартрат холина 0,2; MP смесь витаминная диетическая 1,02; минеральная смесь 3,5; кукурузное масло 8. Состав диеты 40% MetR (г/100 г): L-аргинин 1,124; L-лизин 1,445; L-гистидин 0,331; L-лейцин 1,114; L-изолейцин 0,823; L-валин 0,823; L-треонин 0,823; L-триптофан 0,181; L-метионин 0,516; L-глутаминовая кислота 2,709; L-фенилаланин 1,164; L-глицин 2,338; декстрин 5,017; кукурузный крахмал 31,92; сахароза 31,92; целлюлоза 5,017; битартрат холина 0,201; MP витаминная диетическая 1,023; минеральная смесь 3,512; кукурузное масло 8,028. Продолжительность применения диеты составляла семь недель.
Средняя масса тела животных не показала значимых различий между двумя экспериментальными группами. Значительных изменений потребления кислорода митохондриями не произошло. Все маркеры окислительной модификации митохондриальных белков сердца были достоверно уменьшены на 20–58% в группе MetR. Метилирование геномной ДНК в сердце снизилось с 1,75% до 1,3%. Более сильное понижение наблюдалось у маркера окисления жиров MDAL – 58%, тогда как другие маркеры модификации белка показали снижение на 19–35%. Увеличение продолжительности жизни крыс MetR составило до 50% от таковой у животных на контрольной диете.
Благоприятные эффекты MetR у грызунов также включают снижение концентраций висцерального жира, триглицеридов, холестерина, глюкозы, инсулина и IGF1, замедление развития катаракты, защиту от возрастных изменений иммунитета, улучшение барьерной функции плотного соединения толстой кишки, повышение метаболической гибкости и снижение заболеваемости раком. Сверхэкспрессия метионинсульфоксидредуктазы увеличивает продолжительность жизни животных. Напротив, чрезмерное потребление метионина с пищей оказывает токсические эффекты, включая усиление окислительного стресса. Однако механизм, ответственный за замедление старения с помощью MetR, до сих пор полностью не известен. Поскольку диета снижает окислительное повреждение и продукцию свободных радикалов в митохондриях, можно предположить, что уменьшение окислительного стресса является механизмом увеличения продолжительности жизни при MetR. MetR снижает продукцию митохондриальных АФК в сердце и окислительное повреждение митохондриальной ДНК и белков, и снижает степень метилирования геномной ДНК. Эти результаты показывают, что метионин может быть диетическим фактором, ответственным за снижение выработки митохондриальных АФК и окислительного стресса, а также, вероятно, за частичное увеличения продолжительности жизни [4].
Другие исследования также показали, что диета MetR эффективна для увеличения продолжительности жизни беспородных мышей [5] и крыс с различными возрастными патологиями [6].
Влияние ограничения метионина на биомаркеры при избыточном весе
У мышей восемь недель диеты MetR вызвали 3,1-кратное увеличение чувствительности к инсулину всего тела и увеличение тканеспецифического поглощения глюкозы [7, 8] Кроме того, MetR усиливает стимулированное инсулином фосфорилирование Akt в печени, мышцах, коричневых и белых адипоцитах мышей[8]. Частично из-за сенсибилизирующего к инсулину эффекта MetR можно объяснить его способностью снижать массу тела. Следует отметить, что ограничение метионина эффективно, когда другая заменимая аминокислота – цистеин – отсутствует в диете: включение цистеина отменяет эффекты MetR на метаболизм и антиоксидантный статус [9].
Чтобы исследовать потенциал ограничения метионина и цистеина у людей, в двойном слепом рандомизированном контролируемом пилотном исследовании норвежские ученые оценили влияние диеты на кардиометаболические факторы риска, аминокислотный профиль в плазме и моче, сывороточный фактор роста фибробластов 21 (FGF21) и экспрессию генов подкожной жировой ткани у женщин с избыточным весом и ожирением [10].
Авторы разработали три диеты с различным содержанием серосодержащих аминокислот, регулируя количество добавляемого в рацион порошка смеси метионина и цистеина [10]. Группа, принимавшая добавку с высоким содержанием Met/Cys, считалась контрольной. Средний состав питательных веществ в диете, за исключением порошка Met/Cys, включал жиры 73,4 г, углеводы 264,2 г, белки 86,1 г, насыщенные жирные кислоты (ЖК) 9,5 г, мононенасыщенные ЖК 39,3 г, полиненасыщенные ЖК 17,6 г и по 0,8 г метионина и цистеина; суммарная энергетическая ценность составляла 2190 ккал. Типичное дневное меню было веганское, без мяса, рыбы, яиц, молочных продуктов: на завтрак – овсяная каша с яблоком/изюмом, на обед – салат с фасолью, овощами, фруктами и растительным маслом, на ужин – овощная запеканка/овощной суп/фасолевый салат или овощной суп с хлебом или фокаччой, закуски – орехи или фрукты, и дополнительные протеиновые напитки (с метионином и цистеином или без них) при каждом приеме пищи. Диеты различались только содержанием метионина и цистеина, источником которых были протеиновые напитки. Все диеты соответствовали минимальной рекомендации ВОЗ по суточному потреблению серосодержащих аминокислот. К диете с низким содержанием Met/Cys-low порошок не добавляли; таким образом, среднее содержание серных аминокислот в рационе составляло 1,6 г/день. В диету Met/Cys-medium добавили в общей сложности 2,04 г порошка (0,68 г метионина и 1,36 г цистеина), в результате чего среднее суточное потребление составило 3,64 г день (1,48 г метионина и 2,16 г цистеина). В диету с высоким содержанием Met/Cys-high вводили 4 г порошка (1,32 г метионина и 2,68 г цистеина) с общим средним потреблением 5,6 г/день (2,12 г/день метионина и 3,48 г/день цистеина).
Двадцать женщин с избыточной массой тела или ожирением были распределены в группы с диетой с низким (Met/Cys-low, n = 7), средним (Met/Cys-medium, n = 7) или высоким (Met/Cys-high, n = 6) содержанием метионина и цистеина [10]. Эксперимент продолжался в течение семи дней. Диеты отличались только содержанием метионина и цистеина. Кровь и мочу собирали на 0, 1, 3 и 7 день, а биопсию подкожной жировой ткани брали на 0 и 7 день.
Как и ожидалось, из-за непродолжительности исследования не было убедительных различий в изменении веса между группами. Авторы отметили тенденции к увеличению аполипопротеинов apoА1 и снижению соотношения apoВ к apoА1 и инсулину в группе Met/Cys-low по сравнению с группой Met/Cys-high, но эти результаты были значимыми. Медианные концентрации FGF21 в сыворотке крови увеличились на 47% в группе с низким содержанием Met/Cys от исходного уровня до седьмого дня, тогда как концентрации снизились в группах Met/Cys-medium и Met / Cys-high на 30%, соответственно. Линейный смешанный регрессионный анализ выявил значимое взаимодействие группа × время по ответу FGF21 в группе Met/Cys-low по сравнению с группой Met/Cys-high, но не для группы Met/Cys-medium по сравнению с Met/Cys-high. Для транскриптов, связанных с метаболизмом липидов, наблюдалась повышенная регуляция диацилглицерин-O-ацилтрансферазы 1 (DGAT1), которая участвует в синтезе триацилглицерина, в группе Met/Cys-low с изменением в 0,43log2 раза по сравнению с Met/Cys-high через семь дней. Неубедительные результаты наблюдались для уровней транскрипта ацетил-КоА-карбоксилазы, глутамат-цистеинлигазы, карнитинпальмитоилтрансферазы I и MTOR. Сходные результаты получены для синтазы жирных кислот (FASN). В группе Met/Cys-medium ответ в экспрессии мРНК обычно следовал той же схеме, что и в группе Met/Cys-high [10].
В качестве прокси-маркера активности SCD1 в печени авторы исследовали индекс SCD-16 в плазме (стеароил-КоА-десатураза – фермент, который катализирует образование мононенасыщенных жирных кислот) [10]. Расчетные средние изменения индекса SCD-16 коррелировали со снижением Met/Cys, но результаты были неубедительными. Метионин и цистеин в плазме, а также общий цистеин в моче снизились, тогда как FGF21 увеличился в группе Met/Cys-low по сравнению с Met/Cys-high. Группа Met/Cys-low имела повышенную экспрессию мРНК липогенных генов в жировой ткани, включая DGAT1. Участники сообщили об удовлетворительном соблюдении диеты и умеренной простоте ее соблюдения.
Приведенные данные предполагают, что ограничение метионина и цистеина в рационе может оказывать положительное влияние на циркулирующие биомаркеры, включая FGF21, и влиять на экспрессию генов подкожной жировой ткани [10]. Эти результаты могут быть полезными в разработке и реализации будущих крупномасштабных диетических вмешательств с ограничением метионина и цистеина.
Ограничение метионина и рак
Рак является второй по значимости причиной смерти в Соединенных Штатах: только в 2019 году в США было зарегистрировано 1,7 млн. новых случаев. В глобальном масштабе количество смертей, связанных с раком, превышает смертность, связанную с другими заболеваниями. Незаменимая аминокислота метионин важна для роста и метаболизма раковых клеток. Все больше данных указывает на то, что ограничение метионина подавляет рост онкоклеток и может повысить эффективность химиотерапевтических агентов. Зависимость многих опухолевых клеток от экзогенного источника метионина, делает диетическое ограничение метионина (MetR) потенциальным инструментом в лечении рака. MetR подавляет пролиферацию и рост некоторых типов раковых клеток, тогда как на нормальные клетки ограничение метионина не влияет, по крайней мере, пока присутствует достаточное количество гомоцистеина. В дополнение к ингибированию роста раковых клеток было показано, что MetR увеличивает эффективность химио- и лучевой терапии на животных моделях [11].
Эффект увеличения продолжительности жизни MetR объясняется рядом различных механизмов, включая вызванное MetR снижение окислительного стресса и воспаления, изменение аутофагии и повышение уровня кардиозащитных гормонов. Другой механизм, с помощью которого MetR может влиять на продолжительность жизни, заключается в снижении заболеваемости и общем снижении смертности от рака. Способность MetR улучшать чувствительность к инсулину и уменьшать избыточный вес может быть напрямую связана с его противораковым потенциалом, поскольку существует несколько типов рака, которые тесно связаны с ожирением и инсулинорезистентностью; противораковые эффекты MetR могут быть вторичным по отношению к этим его способностям.
История изучения влияния ограничения метионина на рак началась в 1959 году, когда в одном из первых исследований оценивали эффекты, вызванные диетами, в которых отсутствовали определенные аминокислоты [12]. Эксперимент проводился на крысах, получавших изокалорийную диету, которая содержала все аминокислоты или не содержала одной незаменимой аминокислоты. После трансплантации опухоли Уокера и десятидневной диеты крысы были разделены на разные группы. Каждой группе давали определенную пищу с различным аминокислотным составом в течение пяти дней. Первоначальная цель этого исследования заключалась в том, чтобы различить два противоположных взгляда на азотный баланс и ограничение аминокислот, но результаты показали значительное торможение роста опухоли у крыс, получавших рационы без метионина, валина или изолейцина. Следующее исследование на тканевых культурах было посвящено именно метионину [13]. В линиях клеток рака молочной железы крысы, лимфатической лейкемии мыши или человека, эпителиальных клеток и фибробластах печени крыс, фибробластах кожи мышей, а также клетках рака груди и простаты человека при культивировании в среде, которая либо содержала метионин, либо нет, рост злокачественных клеток был значительно нарушен в составах, обедненных метионином, в то время как рост нормальных клеток не изменился.
С середины 1990-ых годов был проведен ряд исследований ограничения метионина на людях. Так, у четырнадцати человек с раком желудка общее парентеральное питание в течение семи дней до операции с пониженным содержанием метионина, имело синергетический эффект с 5-фторурацилом (500 мг/день; всего 4,0 г/чел) на прогрессирование рака желудка и активность тимидилатсинтазы [14]. Питательный раствор содержал (г/100 мл): L-аргинин 0,66, L-гистидин 0,30, L-изолейцин 0,55, L-лейцин 1,23, L-лизин 1,49, L-фенилаланин 0,87, L-треонин 0,54, L-триптофан 0,18, L-валин 0,61, L-глицин 1,00. Остальные семь из контрольной группы получали 5-фторурацил и обычное парентеральное питание, содержащее 0,71 г/100 мл L-метионина. Общая тимидилатсинтаза снизилась на 9,6% при питании MetR относительно контроля в опухолевых клетках и на 27,4% в нормальных клетках. Гистологический ответ на терапию в группе MetR увеличился как по степени, так и по частоте. Нехватка метионина повышала токсичность 5-фторурацила по отношению к опухолевым клеткам.
Исследование с участием восьми человек с различными метастатическими солидными опухолями (почечно-клеточная карцинома, карциноид, саркома, аденокарцинома поджелудочной железы, аденокарцинома предстательной железы, фолликулярная лимфома) показало, что MetR с энтеральной доставкой снижает концентрацию метионина в плазме и является безопасным и хорошо переносимым для людей с метастатическими солидными опухолями [15].
В исследовании на двадцати пациентах с метастатической меланомой и двух с рецидивирующей глиомой ограничение метионина не показало никаких признаков токсичности или проблем с питанием, но и не оказало влияния на выживаемость [16]. Вес тела был стабильным, как и концентрации сывороточного альбумина и преальбумина.
К настоящему времени количество клинических испытаний ограничения метионина на людях весьма ограничено. Многочисленные примеры действия на признаки рака in vitro и in vivo на животных моделях и клеточных линиях обобщены в обзоре [17].
Ограничение метионина и плотность костей
Как показано выше, ограничение метионина (MetR) увеличивает продолжительность жизни. С другой стороны, обнаружено, что нехватка метионина влияет на плотность костей, врожденный иммунитет и естественные киллеры (клетки NK) [18]. В исследовании мыши 98 дней получали либо базовую диету (BD), содержащую 0,80% метионина, либо пищу с ограничением по метионину, с 0,14% этой аминокислоты. Среднее ежедневное потребление корма было почти одинаковым у мышей BD и MetR в течение всего периода. Однако значительно меньшая масса тела, -25%, была обнаружена у мышей MetR по сравнению с таковой в группе BD через три недели после начала эксперимента, а прибавка в весе была в 17 раз меньше при ограничении метионина. Между тем, уровни общего холестерина, липопротеинов высокой и низкой плотности, и аполипопротеина B (носитель «плохого» холестерина) не изменились в обеих группах. Исследование гормонов, связанных с резистентностью к инсулину, в плазме показало, что мыши с диетой MetR имели повышенный на 57% уровень адипонектина и FGF21 – на 24%, но меньшую концентрацию лептина и IGF-1.
Эксперимент подтвердил опасения, что диета MetR изменяет плотность и структуру костей. Кости мышей из группы MetR были не такими длинными, как в группе BD. Кроме того, наблюдалось снижение минеральной плотности костей на -20% при исследовании с помощью метода DEXA. Животные из группы MetR имели меньшие диаметры медиолатерального и переднезаднего стержней. Как обнаружилось, цитотоксичность NK-клеток была значительно снижена у животных MetR, в отличие от BD, которая имела соотношение клеток 20:1 для мышей MetR и 10:1 у животных BD. Цитотоксичность NK-клеток, рассчитанная по мертвым клеткам-мишеням, при употреблении базовой диеты или с ограничением метионина различалась 4–6%.
Таким образом, мыши из группы MetR демонстрировали снижение костной массы и цитотоксичности NK-клеток селезенки по сравнению с животными BD. Кроме того, мыши с диетой MetR имели меньшую массу тела и более высокие уровни адипонектина и FGF21 в плазме, но пониженные концентрации лептина и IGF-1. В целом, исследование показывает, что нехватка метионина негативно влияет на плотность костной ткани и цитотоксичность NK-клеток. При этом, животные, получавшие MetR-диету, не показали каких-либо других вредных для здоровья побочных эффектов [18].
Заключение
Метионин – незаменимая аминокислота, и поэтому для поддержания жизни ее необходимо употреблять с пищей. Несмотря на то, что потребление метионина необходимо для выживания, исследования показали, что ограничение этой аминокислоты в рационе животных или в средах для культивирования клеток обеспечивает метаболические преимущества, такие как снижение ожирения, повышение чувствительности к инсулину, уменьшая воспаление и окислительного стресса, а также увеличивая продолжительность жизни на животных моделях.
Результаты пилотного испытания с диетами, которые отличались только содержанием метионина и цистеина, показывают, что такое вмешательство возможно для применения на людях и может оказывать эффекты, сравнимые с полученными на животных моделях. Из-за небольшого размера выборки и короткой продолжительности эти результаты следует интерпретировать с осторожностью, но они заслуживают проведения долгосрочных исследований в будущем, чтобы окончательно выяснить, может ли диетическое ограничение метионина и цистеина оказывать устойчивые и положительные эффекты у людей с избыточным весом и ожирением.
Ограничение метионина в пище в течение всего нескольких дней после начала диеты снижает концентрацию циркулирующего метионина в большинстве исследований на людях и животных. Эти данные предполагают, что индуцированное ограничением подавление прогрессирования рака происходит, по крайней мере частично, из-за клеточно-автономных эффектов внутри опухоли. Известно множество связанных с раком изменений в метаболизме с участием метионина, и понимание того, как экзогенный метионин влияет на рак, что может дать важную информацию о новых подходах к лечению. Доклинические исследования показывают, что уменьшение поступления метионина эффективно, когда применяется в контексте профилактики и лечения, и повышает результативность некоторых стандартных химиотерапевтических препаратов против рака. Исследования на моделях in vitro и in vivo показали, что ограничение метионина проявляет противораковую активность при многих типах и стадиях рака.
Ингибирование активности тимидилатсинтазы, снижение биосинтеза полиаминов, индукция апоптоза и изменения метилирования ДНК, образования глутатиона и антиоксидантные эффекты, в зависимости от типа рака, вероятно, являются мишенями при ограничении метионина. Определение механизма, с помощью которого ограничение метионина ингибирует активность тимидилатсинтазы и экспрессию белка, может показать новые лекарственные мишени. Учитывая, что нормальные клетки могут синтезировать достаточное количество метионина для роста, а многим раковым клеткам для выживания требуются значительные количества экзогенного метионина, ограничение метионина имеет некоторый потенциал в качестве терапевтического средства против рака. Выяснение сигнальных путей и молекулярных механизмов ограничения метионина может улучшить существующие терапевтические возможности и привести к разработке новых целевых вариантов лечения для людей, больных раком.
С другой стороны, уменьшение поступления метионина в организм может вести к снижению массы тела, минеральной плотности костей и темпов роста. Потеря веса – симптом, обычно связанный с худшим прогнозом у онкологических больных. Чтобы понять, как повлияют различные варианты ограничения метионина на людей с онкозаболеваниями, необходимы крупномасштабные клинические испытания, сочетающие ограничение метионина в питании с существующими стандартными подходами к лечению онкологических заболеваний.
1. Ables, G.P., Johnson, J.E., Pleiotropic responses to methionine restriction. Experimental Gerontology, 2017. 94: p. 83-88. DOI: https://doi.org/10.1016/j.exger.2017.01.012.
2. Ullmann’s Encyclopedia of Industrial Chemistry. 2011, Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim. p. 29456.
3. Orentreich, N., Matias, J.R., DeFelice, A., Zimmerman, J.A., Low Methionine Ingestion by Rats Extends Life Span. The Journal of Nutrition, 1993. 123(2): p. 269-274. DOI: 10.1093/jn/123.2.269.
4. Sanchez-Roman, I., Gomez, A., Gomez, J., Suarez, H., Sanchez, C., Naudi, A., Ayala, V., Portero-Otin, M., Lopez-Torres, M., Pamplona, R., Barja, G., Forty percent methionine restriction lowers DNA methylation, complex I ROS generation, and oxidative damage to mtDNA and mitochondrial proteins in rat heart. Journal of Bioenergetics and Biomembranes, 2011. 43(6): p. 699-708. DOI: 10.1007/s10863-011-9389-9.
5. Miller, R.A., Buehner, G., Chang, Y., Harper, J.M., Sigler, R., Smith-Wheelock, M., Methionine-deficient diet extends mouse lifespan, slows immune and lens aging, alters glucose, T4, IGF-I and insulin levels, and increases hepatocyte MIF levels and stress resistance. Aging Cell, 2005. 4(3): p. 119-125. DOI: https://doi.org/10.1111/j.1474-9726.2005.00152.x.
6. Zimmerman, J.A., Malloy, V., Krajcik, R., Orentreich, N., Nutritional control of aging. Experimental Gerontology, 2003. 38(1): p. 47-52. DOI: https://doi.org/10.1016/S0531-5565(02)00149-3.
7. Wanders, D., Burk, D.H., Cortez, C.C., Van, N.T., Stone, K.P., Baker, M., Mendoza, T., Mynatt, R.L., Gettys, T.W., UCP1 is an essential mediator of the effects of methionine restriction on energy balance but not insulin sensitivity.The FASEB Journal, 2015. 29(6): p. 2603-2615. DOI: https://doi.org/10.1096/fj.14-270348.
8. Stone, K.P., Wanders, D., Orgeron, M., Cortez, C.C., Gettys, T.W., Mechanisms of Increased In Vivo Insulin Sensitivity by Dietary Methionine Restriction in Mice. Diabetes, 2014. 63(11): p. 3721. DOI: 10.2337/db14-0464.
9. Elshorbagy, A.K., Valdivia-Garcia, M., Mattocks, D.A., Plummer, J.D., Smith, A.D., Drevon, C.A., Refsum, H., Perrone, C.E., Cysteine supplementation reverses methionine restriction effects on rat adiposity: significance of stearoyl-coenzyme A desaturase. J Lipid Res, 2011. 52(1): p. 104-112. DOI: 10.1194/jlr.M010215.
10. Olsen, T., Øvrebø, B., Haj-Yasein, N., Lee, S., Svendsen, K., Hjorth, M., Bastani, N.E., Norheim, F., Drevon, C.A., Refsum, H., Vinknes, K.J., Effects of dietary methionine and cysteine restriction on plasma biomarkers, serum fibroblast growth factor 21, and adipose tissue gene expression in women with overweight or obesity: a double-blind randomized controlled pilot study. Journal of Translational Medicine, 2020. 18(1): p. 122. DOI: 10.1186/s12967-020-02288-x.
11. Gao, X., Sanderson, S.M., Dai, Z., Reid, M.A., Cooper, D.E., Lu, M., Richie, J.P., Jr., Ciccarella, A., Calcagnotto, A., Mikhael, P.G., Mentch, S.J., Liu, J., Ables, G., Kirsch, D.G., Hsu, D.S., Nichenametla, S.N., Locasale, J.W., Dietary methionine influences therapy in mouse cancer models and alters human metabolism. Nature, 2019. 572(7769): p. 397-401. DOI: 10.1038/s41586-019-1437-3.
12. Sugimura, T., Birnbaum, S.M., Winitz, M., Greenstein, J.P., Quantitative nutritional studies with water-soluble, chemically defined diets. VIII. The forced feeding of diets each lacking in one essential amino acid. Archives of Biochemistry and Biophysics, 1959. 81(2): p. 448-455. DOI: https://doi.org/10.1016/0003-9861(59)90225-5.
13. Halpern, B.C., Clark, B.R., Hardy, D.N., Halpern, R.M., Smith, R.A., The Effect of Replacement of Methionine by Homocystine on Survival of Malignant and Normal Adult Mammalian Cells in Culture. Proceedings of the National Academy of Sciences, 1974. 71(4): p. 1133. DOI: 10.1073/pnas.71.4.1133.
14. Goseki, N., Yamazaki, S., Shimojyu, K., Kando, F., Maruyama, M., Endo, M., Koike, M., Takahashi, H., Synergistic Effect of Methionine-depleting Total Parenteral Nutrition with 5-Fluorouracil on Human Gastric Cancer: A Randomized, Prospective Clinical Trial. Japanese Journal of Cancer Research, 1995. 86(5): p. 484-489. DOI: https://doi.org/10.1111/j.1349-7006.1995.tb03082.x.
15. Epner, D.E., Morrow, S., Wilcox, M., Houghton, J.L., Nutrient Intake and Nutritional Indexes in Adults With Metastatic Cancer on a Phase I Clinical Trial of Dietary Methionine Restriction. Nutrition and Cancer, 2002. 42(2): p. 158-166. DOI: 10.1207/S15327914NC422_2.
16. Thivat, E., Farges, M.-C., Bacin, F., Incan, M., Mouret-Reynier, M.-A., Cellarier, E., Madelmont, J.-C., Vasson, M.-P., Chollet, P., Durando, X., Phase II Trial of the Association of a Methionine-free Diet with Cystemustine Therapy in Melanoma and Glioma. Anticancer Research, 2009. 29(12): p. 5235.
17. Wanders, D., Hobson, K., Ji, X., Methionine Restriction and Cancer Biology. Nutrients, 2020. 12(3). DOI: 10.3390/nu12030684.
18. Li, M., Zhai, L., Wei, W., Dong, J., Effect of Methionine Restriction on Bone Density and NK Cell Activity.BioMed Research International, 2016. 2016: p. 3571810. DOI: 10.1155/2016/3571810.




{kind=link}
{kind=link}
{kind=link}
{kind=link}